
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Электрофизиология
Определение "Электрофизиология" в словаре Брокгауза и Ефрона
Электрофизиология
Электрофизиология*
— Э. есть та часть физиологии, которая занимается изучением двоякого рода явлений: 1) электрических явлений, развивающихся самобытно в животном организме, и 2) явлений, происходящих от действия извне электричества на живые существа, их ткани и органы. Первая из этих двух групп явлений состаляет так назыв. животное электричество. Вторая же группа есть ни что иное как совокупность эффектов электрического раздражения мышц, нервов и других тканей организма. Таким образом, Э. распадается на два важные отдела, которые тесно связаны между собой, но тем не менее должны, ради ясности, быть изложены отдельно. Э., начало которой надо искать во второй половине XVIII стол., в темных и запутанных теориях о тождестве нервной силы с электрической, в настоящем своем виде, благодаря тесному общению с физикой и строго научной разработке богатого фактического материала, составляет чуть-ли не самый обширный отдел физиологии. Ни один из других отделов физиологии не пользуется при разработке рассматриваемых им вопросов столь точными и правильными методами исследования, как современная Э.
I. Учение о животном электричестве. — Некоторые ткани животного организма обладают электрическими свойствами, то есть обнаруживают при известных условиях электрические токи, которые могут быть легко определены о помощью электроизмерительных аппаратов. Эти токи наблюдаются, главным образом, в мышцах, нервах и железах, но и другие ткани животного организма и даже все тело животного представляют довольно резкие, хотя еще недостаточно изученные электрические явления. У некоторых рыб, а именно у так назыв. электрических рыб, существуют даже отдельные органы, производящие электрическую энергию. Наконец, у растений наблюдаются тоже весьма характерные электрические явления, находящиеся в связи с их жизнедеятельностью. Таким образом, так называемое животное электричество не есть исключительное свойство животного царства, но составляет общее свойство организованной материи. Собственно говоря, оно должно вернее называться органическим электричеством, под которым надо понимать совокупность явлений животного и растительного электричества. Исторический ход развития учения с животном электричестве составляет, собственно говоря, исторический обзор всей Э. и даже всей электрофизики. Уже с давних пор приписывали электричеству огромное значение в животной жизни и старались даже доказать совершенное тождество нервной силы с электрической. Особенно с точки зрения биологических и философских воззрений XVIII столетия такой взгляд казался совсем естественным и находил до известной степени оправдание в теории электрических разрядов у некоторых рыб, созданной Welsh'eм. Гипотеза отождествления жизненной силы с электрической настолько поглощала тогда внимание всех современных ученых, что даже такие крупные умы, как Гальвани и Вольта, не могли освободиться от этой идеи, влияние которой и обнаруживается в толковании ими фактов первостепенной важности. Открытие лейденской банки не только не поколебало этого мистического учения, но даже в глазах некоторых тогдашних биологов стало еще одним доказательством в пользу электрической природы жизненных явлений. Правду сказать, и теперь еще, в начале XX столетия, некоторые физиологи не теряют надежды путем анализа электрофизиологических явлений открыть тайну природы функции нерва, но как бы ни было велико в этом отношении увлечение этих физиологов, их теории и гипотезы строятся на солидной почве научно установленных фактов, а не на плодах воображения, как это делалось путем чисто спекулятивным в XVIII столетии. Вся Э. того времени и сводится к двум фактам: электрические разряды рыб (скатов) и действие лейденской банки на сокращение мышц животного организма. На этих двух основных фактах и построены все теоретические воззрения на электрическую природу нервной функции. В конце ХVIII столетия Гальвани сделал замечательное открытие, которое пролило яркий свет на вопрос о животном электричестве и сделалось исходной точкой развития всего учения об электричества. Гениальное открытие Гальвани составляет эпоху в науке и должно считаться началом научной Э. Исходный опыт всех электрофизиологических наблюдений Гальвани, оказавшийся столь плодотворным для науки, заключается в следующем. Изучая влияние атмосферного электричества на обнаженные задние конечности лягушки, подвешенные посредством металлических крючков к горизонтальной решетке балкона, Гальвани заметил, что мышцы конечностей вздрагивали всякий раз, когда нижние концы ног, раскачавшись от ветра, приходили в соприкосновение с решеткой. Устранив всякое влияние атмосферного электричества и убедившись в том, что оно совершенно непричастно в данном опыте, гениальный ум Гальвани заключил, что наблюдаемое им явление происходит от замыкания цели, состоящей из мышцы, нерва, металлического крючка и вертикальных стоек металлической решетки. Такая замкнутая цепь может произойти также от соединения лягушечьей лапки со стойкой решетки посредством металлической палочки, как это изображено на фиг. 1.
Фиг. 1. Опыт Гальвани
Повторив этот опыт с прикладыванием металлических дуг к нервно-мышечному препарату и получая всегда мышечное сокращение, Гальвани прибегнул для объяснения этого замечательного явления к чистой гипотезе. Он представил себе мышцу как род лейденской банки, наружная поверхность которой заряжена одним электричеством, а внутренняя — противоположным. Нерв, будучи продолжением внутренней поверхности, составляет кондуктор лейденской банки. Понятно, что при таких условиях прикладывание металлической дуги к нерву и мышце вызывает разряд, который и возбуждает мышцу; отсюда сокращение её. Гальвани предполагал, что у всех животных электричество рождается в мозгу и распространяется отсюда по нервам к мышцам. Эта гипотеза, как бы она ни была невероятна, несомненно указывает на то, что Гальвани искал причину наблюдаемого им явления в "собственном животном электричестве" лягушки и в этом смысле старался истолковать результаты всех опытов, проделанных им впоследствии, с целью пополнить свой исходный опыт. Вольта, проверив опыты Гальвани, считал гипотезу последнего несостоятельной, отрицал существование всякого животного электричества и объяснял наблюдаемое при этих опытах явление как результат соприкосновения металла с животными тканями, или же как эффект соприкосновения двух разнородных металлов между собой. Эти-то опыты и послужили Вольта для открытия его знаменитого "столба" и стали исходной точкой физического учения о гальванизме, но никак не пошатнули веры Гальвани в существование собственной электрогенной силы в животном организме. В начале XIX стол. все внимание ученых было поглощено исследованием физических явлений гальванизма, так что учение о животном электричестве было совсем оставлено в стороне и возродилось только около 1825 г., когда Nobili удалось устроить весьма чувствительный мультипликатор. С помощью этого аппарата Nobili с точностью определил присутствие токов в теле лягушки и их направление от мышц к нервам. Он назвал эти токи собственными токами лягушки. Вскоре после этого Matteucci доказал существование мышечного тока и своими интересными опытами проложил путь для замечательных изысканий du Bois-Reymond'a, составляющих эпоху в науке о животном электричестве. Если Гальвани считается родоначальником учения о животном электричестве, то du Bois-Reymond должен бесспорно считаться основателем всей современной Э. Благодаря его замечательным исследованиям, исполненным по строго научному экспериментальному методу, не только уже раньше известные, но весьма запутанные явления были окончательно разъяснены, и наука обогатилась целым рядом новых фактов, проливших яркий свет на темные вопросы электрогенезиса в животном организме. Дальнейшие исследования других электрофизиологов (Hermann, Hering, Bernstein, Fleischl, Biedermann, Boruttau, d'Arsonval, Jolyet, Burdon-Sanderson, Gotch, Waller, Сеченов, Ламанский, Чирьев, Данилевский, Введенский, Вериго, Мороховец, Мендельсон и др.) составляют только дополнения к работам du Bois-Reymond'a и касаются фактов и теоретических взглядов, которые должны были измениться с развитием науки и с накоплением новых физиологических и физико-химических данных. Заслуга du Bois-Reymond'a заключается еще в том, что он усовершенствовал и изобрел новые электрофизиологические аппараты, основанные на физических началах и создал точные методы исследования, чем и дал возможность следующим поколениям продолжать начатая им изыскания. История Э. за последние 50 лет есть не что иное как изложение фактов, составляющих суть этой науки и тесно связанных с именем du Bois-Reymond'a и его последователей. Благодаря многочисленным работам, сделанным за этот период времени, Э. составляет теперь чуть ли не самый обширный отдел физиологии и представляет громадный интерес не только с теоретической, общебиологической, но и с практической, медицинской точки зрения.
Токи мышц и нервов. Самые наглядные электрические явления наблюдаются в мышцах и нервах, токи которых могут быть отведены к гальванометру и произвести в этом последнем более или менее значительное отклонение магнитной стрелки.
Токи мышц во время покоя. Уже Маттеуччи наблюдал электрические токи в мышцах, но только Дю-Буа-Реймон определил условия их происхождения и формулировал законы, по которым эти токи распространяются. Если поперечный разрез и продольную поверхность любой мышцы лягушки (лучше всего мышцы с параллельно расположенными волокнами, напр., толстой трехглавой мышцы бедра) соединить посредством неполяризуюшихся электродов с гальванометром, то наблюдается в этом последнем более или менее значительное отклонение стрелки, которое указывает на несомненное существование тока в исследуемой мышце. Более того, из направления, в котором отклоняется стрелка, можно заключить, что наблюдаемый ток направляется в мышце от поперечного разреза к её продольной поверхности, следовательно, в гальванометрической цепи, т. е. по проводникам, соединяющим мышцу с гальванометром, ток идет от продольной поверхности мышцы к её поперечному разрезу. Этот ток носит название поперечно-продольного тока мышцы и указывает на то, что поперечный разрез мышцы относится отрицательно к её продольной поверхности, которая наэлектризована положительно. Самый сильный ток идет по направлению от середины поперечного разреза мышцы к её экватору, т. е. к середине её продольной поверхности. В этих двух точках находится максимум электрического потенциала, который убывает по мере удаления от этих точек. Таким образом, более слабые токи получаются от соединения двух точек продольной поверхности, находящихся в неодинаковом расстоянии от экватора или же от соединения двух точек поперечного сечения, неодинаково отстоящих от центра. Две точки, находящиеся на равных расстояниях от середины продольной поверхности мышцы или от центра её поперечного разреза, не представляют разницы электрического потенциала, следовательно, не обнаруживают никакого тока. Совокупность всех этих явлений, уже отчасти известных Matteucci, составляют закон мышечных токов, который был впервые формулирован du Bois-Reymond'ом. Следующая схема показывает силу и направление токов в мышце, снабженной двумя поперечными разрезами и представляющей вид цилиндрической призмы.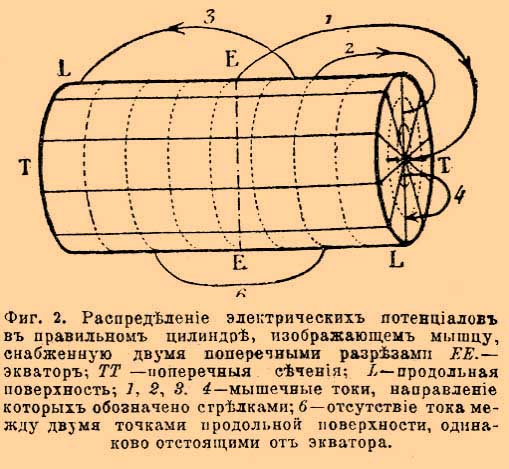
Фиг. 2. Распределение электрических потенциалов в правильном цилиндре, изображающем мышцу, снабженную двумя поперечными разрезами: ЕЕ — экватор; ТТ — поперечные сечения; L — продольная поверхность; 1, 2, 3, 4 — мышечные токи, направление которых обозначено стрелками; 6 — отсутствие тока между двумя точками продольной поверхности, одинаково отстоящими от экватора
Если вместо поперечных разрезов, перпендикулярных к продольной оси мышцы, провести через мышцу два косые поперечные разреза, то вместо цилиндра получится ромб, в котором распределение токов совсем другое, чем в нормальном мышечном цилиндре. Все точки поперечного разреза или продольной поверхности ромба, лежащие ближе к тупому углу, относятся электроположительно ко всем точкам, лежащим ближе к острому углу. Получаемые при таком распределении электрических потенциалов токи называются токами наклонения; они обыкновенно сильнее токов правильного мышечного цилиндра. Некоторые нормальные мышцы имеют тоже ромбоидальную форму в силу особенного расположения своих волокон. Икроножная мышца лягушки представляет даже двойной мышечный ромб; её сухожильная часть, составляющая естественный поперечный разрез мышцы, является электроположительной в отношении к продольной поверхности. Своеобразное направление токов в такой мышце, резко отличающееся от направления токов в нормальной мышце, составляет, по du Bois-Reymond'у, явление парэлектрономии мышц. Достаточно сделать искусственный поперечный разрез, чтобы получить правильное распределение электрического потенциала. Выше описанная законность в распределении электрических токов свойственна не только целой мышце, но наблюдается также в её отдельных кусках. Этот факт указывает на то, что мышечный ток есть явление, присущее каждому отдельному мышечному волокну. Электродвигательная сила мышечного тока равна приблизительно 0,03—0,08 вольта и тесно связана с жизненными свойствами мышцы. Она ослабевает в умирающей мышце и вполне исчезает при наступлении трупного окоченения. В нормальной свежей мышце сила мышечного тока достаточна для того, чтобы раздражать нерв и вызвать сокращение в нервно-мышечном аппарате. Это явление было описано уже давно самим Гальвани под названием: "сокращений без металлов".
Токи в нервах во время покоя. Токи в нервах, открытые впервые du Bois-Reymond'ом (1844), распределяются таким же образом, как и в мышце. Поперечный разрез нерва относится электроотрицательно к его естественной продольной поверхности. Середина продольной поверхности (экватор) нерва и центр его поперечного сечения представляют максимум электрического потенциала, который ослабевает по мере удаления от этих точек. Все токи, наблюдаемые в нерве, сводятся к следующим трем типам: 1) ток поперечно-продольный, идущий от поперечного разреза нерва к его продольной поверхности; 2) ток продольной поверхности получается между двумя точками этой поверхности, лежащими в неодинаковом расстоянии от экватора, и 3) осевой ток, направляющийся от одного поперечного разреза к другому. Каждый из двух поперечных разрезов нерва относится, как было оказано выше, электроотрицательно к продольной поверхности, но оба поперечные разреза, сравниваемые между собой, не представляют одинакового электрического потенциала. В двигательных, т. е. центробежных, нервах поперечный разрез центрального конца является менее электроотрицательным, значит, положительным по отношению к поперечному разрезу периферического конца (du Bois-Reymond). Наоборот, в чувствительных, т. е. центростремительных, нервах периферический поперечный разрез относится положительно к центральному поперечному сечению. Из этого следует, что осевой ток имеет восходящее направление в центробежных (двигательных) нервах и нисходящее направление в центростремительных (чувствительных) нервах, значит, направление осевого тока противоположно направлению физиологической функции нерва (М. Э. Мендельсон). Этот закон осевого тока имеет большой теоретический интерес, так как он прямо указывает на функциональную связь животного электричества с физиологической деятельностью нерва. Направление осевого тока в нерве составляет тоже весьма наглядную и даже единственную разницу между центростремительными и центробежными нервами. Электродвигательная сила нервного тока равна 0,005—0,03 вольта; она зависит от жизненности нерва и от его толщины. Хотя сила нервного тока сравнительно весьма незначительна, но она достаточна для того, чтобы произвести раздражение нерва в другом нервно-мышечном препарате. Замкнув цепь поперечно-продольного тока нерва, можно вызвать так же, как и в мышце, самораздражение данного нерва и, следовательно, сокращение мышцы, иннервируемой этим нервом (du Bois-Reymond, K ü hne, Hering, Biedermann). Если произвести такое же побочное замыкание поперечно-продольного тока в электрическом нерве ската, то можно получить во время замыкания и размыкания тока довольно сильный разряд электрического органа (М. Э. Мендельсон).
Токи действия мышц и нервов. Все вышесказанное относится к покоящимся мышцам и нервам. Но и во время действия мышца и нерв обнаруживают токи, впервые описанные da Bois-Reymond'ом и названные им отрицательным колебанием мышечного и нервного тока. По du Bois-Reymond'y, ток, происходящий во вредя действия мышц и нервов, влечет за собой уменьшение их собственного тока, между тем как по Hermann'y, на эти токи надо смотреть прямо как на токи действия, связанные с процессом возбуждения в мышце и нерве. Если отвести к гальванометру собственный поперечно-продольный ток мышцы или нерва и раздражать эти последние токами, достаточно сильными для того, чтобы произошло мышечное сокращение, то легко можно убедиться в том, что при переходе мышцы или нерва в состояние действия, магнитная стрелка двигается обратно по направлению к нулю. Если она до раздражения остановилась, напр., на 60-м делении шкалы, то в момент мышечного сокращения она может, двигаясь в обратном направлении, остановиться на 30-м или даже на 20-м делении шкалы. Это явление несомненно указывает на то, что собственный ток покоящейся мышцы или нерва значительно ослабел во время их действия и что это ослабление есть результат развития тока обратного направления, составляющего именно отрицательное колебание покойного тока (du Bois-Reymond). Иногда отрицательное колебание может быть столь сильным, что совершенно уничтожает покойный ток; магнитная стрелка возвращается тогда к нулю. Продолжительность отрицательного колебания равна приблизительно 1/300 секунды. Оно распространяется со скоростью 3 метров в секунду (Bernstein). Отрицательное колебание может произвести в другом нервно-мышечном препарате раздражение и сокращение мышцы. Лягушечья лапка, расположенная на бьющемся сердце кролика, раздражается и сокращается от тока действия каждого удара сердца (Маrеу). Во время тетанического, т. е. длительного сокращения мышцы, происходящего от действия известного количества прерывистых раздражений, каждое одиночное раздражение сопровождается соответственным отрицательным колебанием, которое можно легко проследить гальванометрически или электрометрически, или же с помощью телефона (Hermann, d'Arsonval, Введенский). Сила отрицательного колебания в известных границах пропорциональна силе покойного тока. Отрицательное колебание мышечного тока находится в тесной связи с силой сокращения мышцы: в нерве же отрицательное колебание нервного тока служит лучшим и даже единственным средством для обнаруживания деятельности нерва, если нервное возбуждение не сопровождается мышечным сокращением.
Электрические токи в других тканях и органах. — Уже du Bois-Reymond доказал существование электрических токов в коже, наружная поверхность которой является электроположительной в отношении к внутренней поверхности; таким образом ток кожи направляется снаружи внутрь. Подобные электродвигательные свойства представляют тоже разные слизистые оболочки, снабженные железами (Rosenthal, (Gr ü nhagen) и даже кожа рыб, лишенная желез (Hermann). Электрические тока желез обнаруживаются не только при покое, но и во время их действия (секреторные токи). Выделение слюнных желез сопровождается током действия (Bayliss и Bradford). Такой же ток наблюдается и во время секреторной деятельности слизистой оболочки желудка (B iedermann, Bohlen).
Нервные центры также обнаруживают различие электрических потенциалов, хотя токи в спинномозговой оси не представляют того правильного и строго законного распределения, как в мышце и нерве. Это связано, конечно, с весьма сложным строением центральной нервной системы. В продолговатом и спинном мозгу токи покоя и токи действия были впервые наблюдаемы Сеченовым. Такие же токи были определены другими экспериментаторами и в головном мозгу (Caton, В. Я. Данилевский, Fleischl, Beck и Cybulski). Токи действия в этом органе происходят не только от непосредственного возбуждения самого вещества мозга, но и от раздражения любого центростремительного нерва. Передняя половина мозга (лобные доли) относится электроположительно к его задней половине (затылочные доли); но вообще направление токов в головном мозгу весьма разнообразно и не может быть подведено под одну общую схему. Тем не менее законы происхождения и paпределения электрических токов, сопровождающих деятельность головного мозга, составляют несомненно одну из самых важных задач психо-физиологии. К сожалению, вопрос этот еще недостаточно разработан и известные в этом отношении факты не допускают никаких общих заключений. В органах чувств наблюдаются также электродвигательные явления, которые особенно хорошо изучены в глазу. Уже du Bois-Reymond доказал, что зрительный нерв у рыб относится электроотрицательно к передней части (к роговице) глазного яблока. Электрические явления в глазу могут быть вызваны с помощью адекватного, т. е. светового раздражения, причем желтый свет действует сильнее всего в электрогенном смысле, другие цвета действуют слабее (Holmgren, Mac-Kendrick). Токи действия находятся, по всей вероятности, в связи с зрительным пурпуром сетчатки; они значительно ослабевают в сетчатке с обесцвеченным пурпуром (K ü hne, Steiner). Фото-электрические явления могут быть вызваны не только вследствие прямого действия света на исследуемый глаз, но и путем рефлекса, вследствие светового раздражения другого глаза (Engelmann).
Электрические явления у человека составляют одну из самых интересных глав учения о животном электричестве. В начале XVIII столетия, в тот туманный период Э., в котором делались первые попытки применения электрической силы для теоретического объяснения жизненных явлений, мы находим уже целую группу фактов, говорящих в пользу самобытного развития электричества в живом животном организме. Наблюдаемые явления извлечения искр из электризованных людей и животных (Nollet, Gray) заставили многих ученых того времени прямо принять заряжение человеческого тела электричеством и возможность самопроизвольной электризации животных и человека как факт окончательно установленный. Du Bois-Reymond приводит один интересный случай, описанный Озерцовским (1779 г.) на основании рассказов сибиряков, заслуживавших доверия. Некий Михаил Пушкин в Тобольске (1775 г.), стоя зимой на изолирующем ковре, сообщал электрические разряды лицам, прикасавшимся к нему. С того времени и даже в самое последнее время наблюдались многие случаи существования электрогенной способности у невропатических субъектов. Как бы ни были интересны все эти факты, они не могут считаться прямо доказательными в смысле электрогенных свойств человеческого тела уже потому, что при этого рода наблюдениях не всегда исключались с точностью побочные источники для развития электричества, как напр., трение платья о кожу. Ближе к вопросу об электрогенезисе в человеческом теле подошел du Bois-Reymond, изучая прямо электрические токи мыщц при покое и во время действия у человека. Ему не удалось обнаружить у человека ток покоя в мышце при неповрежденной коже, но зато с помощью остроумно придуманного опыта он блистательно доказал развитие электрических токов в мышце под влиянием волевого сокращения мышц. Опыт этот, который изображен на фигуре 3, состоит в следующем.
Фиг. 3. Опыт du Bois-Reymond'а над мышечным током у человека. Ток, направление которого указано стрелками, идет в теле наблюдаемого человека от правого сокращенного плеча к левому несокращенному; следовательно, в гальванической цепи ток идет в обратном направлении
Если погрузить один или несколько пальцев обеих рук в проводящие сосуды гальванометра и сократить мышцы одной руки, то тогда наблюдается отклонение гальванометрической стрелки, указывающее на существование тока, идущего в гальванометрической цепи от расслабленного плеча к сокращенному, следовательно, имеющего в самом человеческом теле обратное направление от сокращенного плеча к расслабленному. Этот ток имеет, таким образом, восходящее направление от руки к плечу. Электродвигательная сила его равна приблизительно 0,002 вольта (Hermann). Эффект получается более наглядный при соединении целого ряда лиц, которые держат друг друга за руку и одновременно сокращают одноименную конечность. Hermann считает эти токи секреторными токами кожи, но многие факты говорят в пользу того, что наблюдаемые du Bois-Reymond'ом токи у человека под влиянием волевого сокращения мышц суть нечто иное как токи действия, сопровождающие деятельность мышц. Они наблюдаются и у таких субъектов, у которых секреторная дятельность кожи совсем прекращена, вследствие патологических изменений в коже и подвергаются различным уклонениям при разных заболеваниях нервно-мышечной системы (М. Э. Мендельсон). По Waller'y сердце у человека обнаруживает также токи действия, которые сопровождаются изменениями электрического потенциала на поверхности всего тела. Тарханов наблюдал даже развитие секреторных токов в коже у человека во время психической деятельности и под влиянием разных волнений. Вероятно, деятельность мног. других тканей и органов человеческого организма сопровождается тоже электрическими явлениями, присутствие которых еще не обнаружено и изучение которых составляет ближайшую задачу современной Э. Несомненно вся органическая природа снабжена электрогенными свойствами, которые проявляются только при известных условиях. У разных животных, у кошек, у крыс и даже у лошадей (Dehmer) были наблюдаемы явления животного электричества. У птиц перья заряжаются при взмахивании положительным электричеством в отношении к воздуху, который электронегативен. Но самые замечательные электрические явления наблюдаются у некоторых рыб (Torpedo, Gymnotus, Malepterurus), снабженных специальным электрическим органом (см. ст. Электрические рыбы). Явления электрогенезиса присущи не только животному царству, а наблюдаются также и в растительном царстве. Электрические токи определенного направления сопровождают пассивные и активные движения растений (Munk, Hermann, Burdon-Sanderson, Kunkel); они особенно резко выражены у раздражительных (напр., у Mimosa pudica) и у мясоядных растений, как у Dionae muscipula. Сгибание листьев у этих растений вследствие искусственных или естественных раздражений сопровождается развитием электрических токов, представляющих известную аналогию с отрицательным колебанием мышечного тока.
Tеории животного электричества. Весьма естественно, что блистательные открытия du Bois-Reymond'a в области животного электричества заставили его искать объяснение этих явлений и создать теорию, составляющую логическое последствие всех наблюдаемых им опытных данных и известную под названием молекулярной meopиu du Bois-Reymond'a. В силу этой теории каждое мышечное или нервное волокно состоит из бесконечного числа мельчайших электродвигательных, периполярных молекул, погруженных в индифферентной проводящей жидкости и имеющих один положительный экваториальный пояс, обращенный к поверхности и два полярных пояса отрицательных, обращенных к поперечным разрезам. Эта физическая электромолекулярная схема объясняет более или менее удовлетворительно происхождение различных токов покоя и действия в мышцах и нервах. Основной принцип этой теории состоит в том, что электрические явления в мышцах, нервах и других животных тканях представляют продукт их живой организации; они существуют во время покоя так же, как и во время действия, и тесно связаны с жизненными свойствами организма. Этот принцип предсуществования (Preexistenz) электродвигательных свойств животных тканей сильно оспаривается L. Hermann'ом, основателем другой теории животного электричества, известной в науке под названием альтерацuoннoй meopиu (Alterationsteone). По Hermann'у, электрические токи не предсуществуют в неповрежденных нервах или мышцах; они являются последствием повреждения, причиненного поперечным сечением мышцы или нерва, причем поврежденная (демаркационная) поверхность как умирающее вещество является электроотрицательной по отношению к остальной неповрежденной части нервно-мышечной ткани. Раздраженное место нерва или мышцы относится тоже электроотрицательно к нераздраженному месту. Таким образом и объясняется происхождение токов покоя (демаркационных токов) и токов действия. В противоположность теории du Bois-Reymond'a, которую можно считать физической, теория Hermann'a вся основана на химических началах. Becquerel и вслед за ним d'A r sonval старались свести явления животного электричества к так называемым электрокапиллярным явлениям, играющим бесспорно весьма важную роль в жизненных отправлениях животного организма. Но ни одна из этих теорий не объясняет вполне и с достаточной точностью электрогенные свойства живой материи. Надо надеяться, что новые данные электрохимии вскоре дадут более удовлетворительное объяснение электрогенезиса в организованной природе. Но пока можно сказать, что из всех упомянутых теорий электромолекулярная теория du Bois-Reymond'a ближе всего подходит к истине, особенно если ее видоизменить согласно новым данным современной электрохимии.
II. Явления, происходящие от действия электричества на нервы и мышцы. Этот отдел Э. не представляет того общебиологического интереса, какое имеет учение о животном электричестве, затрагивающее самые жгучие вопросы жизни, но, тем не менее, он интересен в том отношении, что знакомит с механизмом функциональной деятельности мышц и нервов. Нервная и мышечная ткань обладают свойством раздражительности, в силу которого эти ткани под влиянием какого-нибудь раздражения переходят в деятельное состояние, аналогичное тому физиологическому возбуждению, которое происходит от действия естественного возбудителя, т. е. нервного импульса, исходящего из нервных центров. Это особое деятельное состояние (состояние возбуждения) выражается мышечным сокращением в мышце и током действия в нерве. Впрочем, возбуждение двигательного нерва, находящегося в связи с мышцей, проявляется тоже в виде мышечного сокращения, сила которого указывает на энергию возбуждения в нерве. Из всех искусственных раздражителей электрический ток составляет самый точный и более всего приближающийся к естественному раздражитель. Поэтому он и употребляется чаще всего при физиологических опытах. Для раздражения нервов и мышц употребляются разные виды электрической энергии, но более всего изучено до сих пор действие гальванических и индукционных токов. Что касается вообще раздражающего действия электрического тока на нервно-мышечный аппарат, то оно подчинено следующему закону, впервые формулированному du Bois-Reymond'ом: не абсолютная сила тока, а колебания этой силы производят в нерве или мышце раздражающее действие. Покуда сила тока остается равномерной и ток циркулирует без всяких колебаний, не происходит никакого эффекта возбуждение в нервно-мышечном аппарате; деятельность этого последнего обнаруживается только при внезапном изменении силы тока или в моменты замыкания (возникновения) и размыкания (исчезания) тока. В промежутках между этими моментами электрический ток не производит никакого видимого раздражающего действия. Чем быстрее происходят колебания силы тока, тем эффект электрического раздражения сильнее. Но все-таки быстрота колебаний не должна перейти известных границ, и продолжительность электрического тока должна быть не менее 0,0015 сек. для того, чтобы произошло возбуждение нерва. Вообще нерв отличается большей раздражительностью по отношению к электрическим раздражителям, чем мышца. Нервы у новорожденных менее возбудимы, чем у взрослых. Эффект электрического раздражения зависит от длины раздражаемого нерва от силы и от направления тока. Продольные токи, т. е. идущие вдоль нерва, несравненно действительнее поперечных токов. Различные точки нерва неодинаково чувствительны к электрическому току; точки, лежащие ближе к центру, более возбудимы, чем точки, находящиеся ближе к периферии. Если нервно-мышечный аппарат раздражать отдельными короткими колебаниями (ударами) тока, весьма быстро следующими друг за другом, то вместо одиночного вздрагивания мышцы, соответствующего одиночному раздражению, получится более или менее продолжительный столбняк, сила и продолжительность которого находятся в известных границах, в прямом отношении к силе и частоте отдельных раздражений. Впрочем, для токов максимальной силы существует optimum и pessimum частоты, для токов же менее частых существует optimum и pessimum интенсивности (Введенский). Действие постоянного гальванического тока не ограничивается лишь эффектами раздражения, а производит при прохождении через нерв и мышцу весьма характерные изменения возбудимости и электродвигательных свойств, известные под названием электротона. Этими-то электротоническими изменениями нерва и мышцы объясняется закон сокращений, основанный на полярности действия электрического тока (см. Электротон). Гальванический и вообще электрический ток производит еще на многих низших и некоторых высших животных так называемое направляющее действие (galvanotaxia), в силу которого животные распределяются своеобразно между электродами и даже двигаются к определенному полюсу (см. Электротаксис). Большинство описанных выше явлений наблюдается одинаково при действии гальванического и индукционного тока, так как действие этого последнего весьма близко подходит к действию прерывистых гальванических токов. При равной силе индуцирующего тока размыкательные индукционные удары действуют сильнее замыкательных. Действие статического электричества и разрядов конденсатора на нервы и мышцы в общем не отличается от действия мгновенных постоянных токов. Физиологическое действие токов высокого напряжения и большой частоты на животный организм еще недостаточно изучено. По исследованиям d'Arsonval'я, эти токи имеют весьма благоприятное влияние на обмен веществ и общее питание организма, но это действие оспаривается многими другими исследователями. В самое последнее время В. Я. Данилевский изучал физиологическое действие электричества на расстоянии, с помощью так называемого электро-кинетического раздражения нервов, т. е. беспосредственного воздействия электричества через диэлектрик без участия каких-либо межуточных проводников. Полученная при этом "физиологическая реакция" является результатом известного электрического процесса, индуктивно вызванного в самом организованном живом объекте (В. Данилевский).
Литература. Электрофизиологическая литература столь обширна, что нет возможности дать ее здесь целиком. Желающие ближе познакомиться с затронутыми выше вопросами найдут их обстоятельное изложение и всю литературу до последнего времени в следующих работах и руководствах: Galvani, "De viribus electricitatis in motu musculari commentarius" (Болонья, 1791); его же, "Dell'uso et dell'attivita dell'arco conduttore nelle contrazioni dei musculi" (Болонья, 1794); Matteucci, "Essai sur les ph énomènes électriques des a nimaux" (П., 1840); его же, "Trait é des phénomè nes electro-physiologiques des animaux" (Пар., 1844); его же, "Cours d'electro-physiologie" (Пар., 1858); E. Du Bois-Reymond, "Untersuchungen über tierische Electricitä t" (1844 — 48); его же; "Gesammelte Abhandlungen zur Allgemeinen Muskel- und Nerven-physik" (1875—77); И. Сеченов, "О животном электричестве" (1862); L. Hermann, "Handbuch der Physiologie" (т. I, и II ч. 1-я, 1879); W. Biedermann, "Electrophysiologie" (Йена, 1895); его же, "Electrophysiologie" (в Ascher u. Spiro "Ergebnisse der Physiologie", 1902 и 1903); M. Э. Мендельсон, "Electricit é animale" и "Electrotonus" в "Dictionnaire de Physiologie de Ch. Richet" (т. V, 1900); его же,
| "БРОКГАУЗ И ЕФРОН" >> "Э" >> "ЭЛ" >> "ЭЛЕ" >> "ЭЛЕК" |
Статья про "Электрофизиология" в словаре Брокгауза и Ефрона была прочитана 2119 раз
| Пицца в сковороде |
| Луковый соус |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел