
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Экспериментальная эмбриология
Определение "Экспериментальная эмбриология" в словаре Брокгауза и Ефрона
Экспериментальная эмбриология
Экспериментальная эмбриология*
иначе механика развития, хотя было бы правильнее ее называть физиологией развития — занимается изучением развития животных при искусственных условиях. Если эмбриология (см.) вообще изучает морфологическую сторону развития и пытается ответить на вопрос, как оно происходит, то Э. эмбриология изучает физиологическую сторону развития и пытается ответить на вопрос, почему развитие идет так, а не иначе. Конечно, относительно каждого эмбриологического опыта можно возразить, что получение того или другого результата при искусственных условиях не есть еще доказательство, что этот результат, наблюдаемый при нормальных условиях, непременно вызывается той же причиной, как и в нашем опыте. Причины могут быть различны, а результат — одинаков. Но в таких сложных явлениях, как эмбриональные, даже достижение некоторой аналогии с природой является громадным шагом вперед. Собственно, попытки Э. изучения эмбриологии делались давно, но они ограничивались тесной областью изучения условий возникновения уродств и аномалий у цыпленка (см. Тератология). Первый Геккель пробовал удалить части развивающегося яйца сифонофор. Но настоящее обоснование Э. эмбриология получила благодаря трудам Ру (Rоих), а вслед за ним и целого ряда исследователей. Э. эмбриология слишком молода, чтобы можно было дать исторический очерк ее развития. Наметим лишь направления, в которых производились эксперименты. Отметим, что тератология под влиянием Э. эмбриологии получила особое значение: каждое уродство является, с этой точки зрения, не чем иным, как опытом, произведенным самой природой. Изучается в настоящее время влияние силы тяжести при помощи центробежной машины (Пфлюгер, Ру, Борн, Шульце, Московский), влияние температуры (Дриш, О. Гертвиг, Шульце), влияние света (Дриш, Леб), влияние нарушения газового обмена при перенесении яиц в ненормальную атмосферу (Леб, Самасса, Галле); влияние растворов солей (Гербст, Гурвич, Гертвиг, Леб, Морган, Шимкевич), влияние электрического тока (Ру, Росси), магнетизма, сотрясения и т. п. Весьма интересные результаты получены при изучении результатов сдавливания дробящегося яйца (Пфлюгер, Гертвиг, Ру, Борн, Дриш, Циглер) и удаления частей яйца (Ру, Шабри, Барфурт, Дриш, Фишель, Гертвиг, Герлицка, Уильсон, Морган, Эндрес и др.). Конечно, мы не имеем возможности излагать все полученные результаты, но можем наметить главнейшие задачи и результаты, достигнутые Э. эмбриологией. Начнем с влияния силы тяжести. Для многих случаев развития двусимметричных животных доказано, что первая борозда дробления совпадает с срединной плоскостью будущего зародыша, делящей его на правую и левую стороны. Мы можем в богатых желтком яйцах изменить положение первой борозды дробления, при чем изменится положение и срединной плоскости зародыша. Лягушечье яйцо имеет всегда одну сторону темную, пигментированную и обращенную, плавая в воде, вверх. В этой темной части яйца лежит ядро, или зародышевый пузырек, с окружающей его протоплазмой, тогда как на нижнем полюсе сосредоточен желток, как более тяжелая часть яйца. При нормальном развитии первая меридиональная борозда делит темную и светлую части яйца пополам. Но если мы искусственно выведем яйцо из этого положения, так чтобы темный полюс лежал не на верху, а где-нибудь сбоку (фиг. 1), то под влиянием силы тяжести желток постепенно будет перемещаться на нижний полюс, а зародышевый пузырек с окружающей протоплазмой будет перемещаться к верхнему полюсу изменившего свое положение яйца.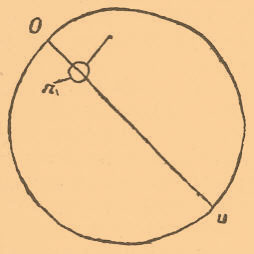
Фиг. I. Схема яйца амфибии, выведенного из нормального положения. o — верхний полюс, u — нижний; линия, соединяющая их, — ось яйца, n — ядро; стрелка указывает направление движения ядра при перемещении его.
Тот же результат может быть достигнут, конечно, если заменить действие силы тяжести влиянием вращения на центробежной машине. Но с перемещением ядра переместится и первая борозда дробления, а расположение пигмента не изменяется. Эта первая борозда, образующаяся, конечно, за разделением ядра на две части, будет делить пигментированную и непигментированную части не пополам, а под некоторым углом, отрезая от пигментированной части меньший участок для одной бластомеры, чем для другой. Итак, первая борозда дробления делит яйцо сообразно с положением ядра и желтка, а смещение желтка влияет на положение первой борозды (Pfl ü ger, 1883—1884; Born, 1893). Однако при опытах над яйцами, бедными желтком, напр. над яйцами морских ежей, такого влияния не замечено (Hertwig, 1884). Вообще же первая борозда в круглых яйцах, по-видимому, не имеет определенного положения, а в яйцах овальных — совпадает с продольным диаметром (Hertwig, 1893). Совпадение это не является вполне строгим, а продольная плоскость зародыша образует с первой плоскостью дробления некоторый угол, коего величина колеблется в известных пределах. Последующие плоскости дробления не имеют определенного отношения к осям зародыша, определяющимся лишь после гаструляции (Kopsch, 1900). Во всяком случае мы можем говорить, имея в виду лишь большинство случаев, а не все. Иногда с продольной осью зародыша совпадает вторая плоскость дробления, иногда не совпадает ни одна, и, наконец, иногда (Ascaris и круглые черви вообще) первая плоскость проходит по экватору яйца и является горизонтальной по отношению к зародышу. Явления дробления сами по себе так сложны, что мы можем выводить не законы, а лишь общие правила, допускающие иногда многочисленные исключения. Однако вопрос относительно значения силы тяжести при нормальных условиях развития весьма сложен. Если первая борозда определяет иногда положение будущего зародыша, то возникает вопрос, чем она определяется сама. Ру полагает, что первая борозда проходит по той же плоскости, по которой проходит живчик к ядру. У амфибий путь этот обозначается полоской пигмента, увлекаемого из периферических слоев яйца живчиком при своем прохождении. Но в то же время мы видели, что существует зависимость этой борозды от влияния силы тяжести, т. е. от распределения желтка. Сила тяжести влияет, конечно, и на яйца, находящиеся в нормальных условиях. Возникает вопрос, почему эти два фактора совпадают. Московский полагает, что совпадение влияния этих двух факторов может быть объяснено только зависимостью их от какого-нибудь третьего фактора, и таковым он считает протоплазматические токи, возникающие в яйце. Яйцо амфибий может свободно вращаться внутри своих оболочек, но в то время, когда проникает в него живчик, означенное вращение невозможно и яйцо остается как бы фиксированным. Яйцо реагирует в этот момент возникающими в нем протоплазматическими токами, а эти последние определяют, с одной стороны, путь прохождения живчика, а с другой — распределение желтка. Оба эти фактора в свою очередь определяют положение первой борозды дробления. Момент вхождения живчика настолько важен в этом отношении, что он определяет собой дальнейший ход, и если бы потом яйцо и подвергалось вращению, то это последнее не может изменить влияния указанного момента. Точка зрения Московского важна потому, что позволяет примирить противоположные взгляды на значение силы тяжести для нормального развития. Одни (Ру, Московский) доказывают необходимость этой силы для нормального развития яиц, богатых желтком; другие (Шульце, Морган, Катаринер) отрицают ее необходимость. Морган, например, помещал яйца амфибий в сосуд, вода коего находилась в постоянном движении, и так как при этом получились нормальные зародыши, то он сделал заключение, что сила тяжести не оказывает влияния на нормальный ход развития. С точки зрения Московского, опыт этот объясняется тем, что сила тяжести влияет лишь в некоторый определенный момент, после которого это влияние действительно может быть устранено без ущерба для нормального развития. Действием центробежной машины Гертвиг обратил лягушечье яйцо, проходящее полное и неравномерное дробление, в яйцо с частичным дроблением. При вращении яйца на центробежной машине ядра, образующиеся делением ядра яйца, остаются около анимального полюса, и только эта часть яйца дробится, тогда как прочая часть остается нераздробленной, хотя в ней после образования около анимального полюса сегментационной полости замечаются в поверхностных слоях желтка элементы, напоминающие собой мероциты меробластических яиц. Весьма может быть, что при действии центробежной машины происходит некоторое уплотнение желтка и такой уплотненный желток оказывает большее сопротивление дробящему влиянию активной протоплазмы, чем нормальный, почему и остается нераздробленным. Такие зародыши могут потом, после прекращения действия центробежной машины, развиваться дальше, но часто дают уродов, а именно таких, у которых обе половины нервной трубки и всей спинной стороны являются не сросшимися, а разделенными вдоль (Spina bifida). Интересные данные получены касательно влияния давления на яйца и на клетки вообще. Деление клетки, а также и яйца может быть замедлено давлением, погружением в растворы солей, недостаточным притоком воды и вообще уплотнением протоплазмы, но это не останавливает деления ядра, которое при возвращении яйца к нормальным условиям иногда сразу вызывает разделение яйца на столько участков, на сколько разделилось ядро. На направление борозд дробления едва ли не самое главное влияние оказывают условия давления на яйцо. Если мы сдавим яйцо лягушки или тритона сверху вниз, то после 2-х меридиональных делений, происходящих как и при нормальных условиях, следует не горизонтальное деление, которое при нормальных условиях приводит к расположению 8-и шаров дробления в два этажа, а опять меридиональное, приводящее к расположению 8-и шаров в один слой в виде пластинки (фиг. 2, А).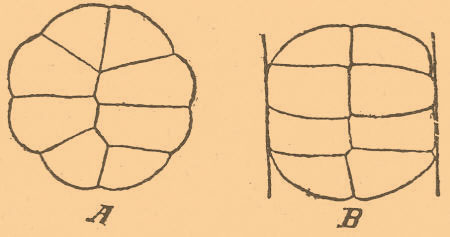
Фиг. 2. Ненормально дробящиеся яйца лягушки. А — между двумя горизонтальными стеклянными пластинками (при рассматривании сверху); В — сдавленные в стеклянной трубочке (по О. Гертвигу из Берга).
Точно так же если ввести яйцо амфибии в трубку более узкого диаметра, чем само яйцо, то оно будет делиться главным образом, хотя не исключительно, в направлении, поперечном к длинной оси трубки (фиг. 2, В). Таким образом, если мы сдавим яйцо с двух сторон, то оно разделится бороздой, перпендикулярной давящим плоскостям, и, следовательно, ядро дробления разделилось в направлении, перпендикулярном к этой борозде, т. е. параллельно давящим плоскостям. Подобные опыты были произведены многими исследователями над яйцами амфибий и морских ежей, причем иногда получалось расположение шаров в виде однослойной пластинки и в более поздних стадиях, с большим числом шаров (Pfl ü ger, 1883—84; Hertwig и Born, 1893; Driesch, 1892; Ziegler, 1894 и др.). Но формулировать результаты этих опытов не так легко. По Гертвигу, вытянувшееся в виде веретена ядро при этом становится параллельно наибольшей оси клетки; по Пфлюгеру — деление ядра происходит в том направлении, где оно встречает наименьшее сопротивление; по Бертольду — клетки делятся по плоскости своего наименьшего сечения. Таким образом пытались формулировать полученные результаты, причем каждая из этих формулировок обнимает лишь известную совокупность случаев, отчасти совпадая с прочими. Если мы, например, примем формулировку Гертвига или Пфлюгера, то нам станет непонятно деление клеток в однослойном эпителии. Клетки перед делением округляются, и, следовательно, все оси их равны, а сдавливаются они со всех сторон, кроме одной — наружной, следовательно, наибольшее давление испытывают с боков. Точка зрения Гертвига неприменима, ибо все оси клетки равны, а при взгляде Пфлюгера мы должны были бы ожидать, что их ядра будут всегда делиться в направлении, перпендикулярном к поверхности эпителия, ибо с наружной поверхности ядро не встречает сопротивления (Heidenhain, 1896). Но этого никогда не бывает, и ядра однослойного эпителия клеток делятся всегда в плоскости, параллельной к поверхности эпителия. Поэтому Брэм (Вraem, 1894) предложил такое правило: клетка делится так, что расходящиеся части ядра встречают не только наименьшее, но и равномерное сопротивление. При разделении эпителиальной клетки в направлении, перпендикулярном к поверхности, наружная часть ядра встречала бы гораздо меньшее сопротивление, чем внутренняя. Вообще, мы покуда можем говорить лишь о некоторых правилах, а не о законах, ибо исключения из этих правил встречаются нередко. Морган (1896) наблюдал такие исключения при дроблении обрывков яиц морского ежа, Дженнингс (1896) многочисленные уклонения наблюдал при дроблении яйца коловратки Asplanchna и т. п. Очевидно, что, кроме условий давления, есть еще иные условия, а может быть, и свойства, присущие самой клетке и определяющие иногда направление деления. Гертвиг думал, что ему, изменяя давлением форму дробления, удалось вызвать такое изменение в расположении шаров дробления, которое изменяло и роль этих шаров в истории развития. Если при нормальном дроблении третья борозда делит яйцо на две части, верхнюю и нижнюю, которые соответствуют спинной и брюшной стороне зародыша, то при сдавливании в горизонтальном направлении эта борозда заменяется меридиональными бороздами и те шары, которые должны были образовать спинную сторону зародыша, ложатся рядом с теми, которые должны образовать брюшную. Следовательно, судьба этих шаров будет иная. Освободим яйцо от давления, и оно, несмотря на ненормальное положение шаров, даст вполне нормальный зародыш, а следовательно, изменение в расположении шаров осталось без влияния на ход развития. Иначе говоря, все шары дробления являются между собой разнозначащими. Но опыты эти допускают и другое толкование. Давление, изменяя форму дробления, не изменяет его, по-видимому, сущности (Heider, 1897). Оно вызывает лишь гетерохронию, или иной порядок во время появления отдельных борозд. При нормальном давлении последовательность деления такая: 1-е меридиональное деление, 2-е — меридиональное, 3-е — экваториальное, 4-е — меридиональное. В результате получается яйцо из 8-и шаров верхних и 8-и нижних. При давлении сверху вниз последовательность такая: 1, 2 и 3-е — меридиональные деления, а затем следует все-таки 4-е горизонтальное и тот же результат, что и в первом случае. В других случаях горизонтальное деление еще более запаздывает, но все же оно неизбежно появляется. Очевидно, что в разбираемом примере 3 и 4 деления только поменялись своей очередью. Опыты Гертвига и других, таким образом, не решают вопроса о том, разнозначащи ли шары дробления или нет. Более убедительны данные, получаемые путем (см. ниже) удаления частей яйца. Впрочем, Дриш утверждает, что ему удалось согреванием и встряхиванием яиц морского ежа, а также ланцетника производить смещение бластомер до полной неправильности и что такие яйца все-таки давали нормальные бластулы и гаструлы. Маас вызывал смещение бластомер у одной медузы, так что они располагались в один длинный ряд, а потом снова располагались сферически и получался нормальный зародыш. Интересно, что у турбелларий, по Галле, у сосальщика Polystomum, по Halkin, а равно, по Мечникову, у одной из медуз (Oceania armata) при нормальном развитии бластомеры располагаются совершенно неправильно и в беспорядке. Давление влияет и на гаструляцию. Так, у Chondracanthus нормально погружается внутрь для образования энтодермы двойной ряд симметрично расположенных клеток, а в яйцах, сдавленных с боков, погружается лишь один ряд, который потом, по-видимому, все-таки делается двойным (Шимкевич, 1895). Что касается до влияния электрических токов, то Росси (1896) заметил, что постоянный ток влияет на направление плоскостей дробления, вызывая некоторые неправильности. Вообще же под влиянием тока клетки (эпидермиса головастика) и ядра в месте приложения электродов могут вытягиваться по направлению тока, но на самое направление деления ток влияния не оказывает (Galeotti, 1896). Если световые, например, условия не влияют на первые стадии, то с началом процесса усвоения желтка в некоторых случаях может быть обнаружено влияние световых и цветных лучей. Яйца саламандры в стадии образования нервной трубки умирали в темноте, хотя в начальных стадиях переносили ее без вреда. Прежде всего темнота влияет подавляющим образом на развитие пигмента (Chiaruugi e Livini 1897; List, 1899), но и то не всегда: некоторые пещерные формы, как змея Rinophis и некоторые турбелларии, являются пигментированными. Наконец, у млекопитающих развивающихся в теле матери, пигмент может появляться во время утробной жизни. Задерживающее действие пониженной температуры было известно давно. О. Гертвиг произвел более точные опыты над яйцами морских ежей и показал, что при действии температуры ниже —1 до —4 в течение 15—30 минут деление клеток останавливается, ахроматинная фигура исчезает, но расположение хроматина остается таким, каким оно было в начале опыта. При более продолжительном действии холода хромосомы могут сливаться или принимать форму пузырьков. Но с возвращением к нормальной температуре деление снова возобновляется. Вообще же, конечно, процесс размножения клеток, как и всякий физиологический процесс, имеет свой optimum, но этот optimum различен даже у близких. Температуры за пределами минимальной и максимальной вызывают или остановку в развитии, или уродливые уклонения. При 2—3° яйца амфибий дают аномальные уклонения, а при 0° останавливаются вовсе в развитии. Впрочем, вопрос о предельной температуре для нормального развития лягушечьего яйца является спорным. По Гертвигу, при 0° происходит остановка, а по Шульце (1899), при 0° развитие хотя очень медленно, но продолжается. Через 30 дней яйцо находится еще в стадии гаструлы. Шульце утверждает, что яйца лягушки, переносившие и боле низкую температуру в течение 14 дней, будучи перенесены в комнатную температуру, давали нормальных зародышей. Вообще, изменение температурных условий является одним из наиболее удобных приемов для получения уклонений (см. Уродства). Повышенная температура вызывает расхождение бластомер и образование двойников (Дриш), появление экзогаструл (см.) и т. п. Другой замедляющий или ускоряющий деление фактор — это газовая среда, причем здесь можно видеть различное отношение к внешним влияниям протоплазмы и ядра. Отсутствие кислорода оказывает обыкновенно замедляющее действие на деление, но при этом наблюдаются, однако, и иные явления: яйца одних рыб (Ctenolabrus), морских ежей в отсутствие кислорода не подвергаются делению, но яйца другой рыбы (Fundulus) — делятся в течение 12 часов (Loeb, 1896), а яйца аскарид, по Галле (1895), остаются живыми в угольной кислоте 4 недели. Многое зависит, впрочем, от того, в какой стадии находится яйцо. Яйца лягушки в атмосфере водорода развиваются в первых стадиях, но в более поздних (в стадии blastula) — погибают (Самасса, 1 8 96), ибо более развитые яйца более нуждаются в кислороде. Впрочем, Шульце получил (1899) несколько иной результат над яйцами лягушки: в отсутствие кислорода они останавливались в развитии, но при возобновлении вентиляции даже через 2 дня развитие возобновлялось, хотя в большинстве случаев результатом его были ненормальные уклонения или яйцо умирало. Кислород, по Демоору, экспериментировавшему главным образом над водорослями, будто бы ускоряет деление, тогда как в опытах других наблюдателей, именно на яйцах лягушки (Самасса) — увеличение содержания кислорода не влияло на скорость деления. Таким образом, из этих не вполне согласных данных мы можем заключить, что влияние газов, вероятно, стоит в большой зависимости от природных свойств клетки, причем эти свойства в различные фазы жизни организма могут подвергаться колебаниям и даже полному изменению. В общих чертах можно сказать, что по мере хода развития яйца потребность в кислороде возрастает. Что касается до действия растворов, то многими было замечено, что погружение яйца в соляной раствор или усиление концентрации морской воды действует замедляющим образом на деление клетки, но и здесь было замечено, что иногда протоплазма останавливается в делении, а ядро продолжает делиться, как это следует из наблюдений Деба и Моргана над яйцами морских ежей. Такое же замедляющее действие было обнаружено при опытах со многими другими солями, причем главным фактором, по-видимому, является не химическое действие того или другого вещества, а нарушение и изменение осмотических условий, вызываемое раствором. Поваренная соль действует вообще как водоотнимающее средство, а этим она сгущает протоплазму и уменьшает ее раздражимость, и следовательно, затрудняет ее деление. Вообще, сжатие яйца или недостаточный приток воды (Циглер, 1 8 94—95) останавливают деление протоплазмы, хотя деление ядра может и совершаться при этом. Фере произвел целый ряд опытов над действием весьма разнообразных веществ на куриное яйцо, и в то время, как одни, напр. алкоголь, оказывали замедляющее действие, другие, напр. никотин, — ускоряющее. Однако эти данные нуждаются в поверке. Так, напр., ускоряющее действие никотина не подтверждается (Шимкевич, 1902). По Лебу (1898), слабые щелочи влияют ускоряющим образом на рост и развитие личинок морских ежей, а кислоты — замедляющим (относительно образования в растворах искусственных лучистостей см. Центросфера). Растворы некоторых солей при действии их на яйца в самом начале развития влияют весьма различно и подавляют то одни, то другие зачатки (Hertwig, 1895, 97; Wil s on, 1897; Gurwitsch, 1896). Так, в различных случаях то не замыкается бластопор, то не срастаются края спинномозговой трубки, то не замыкается мозговой пузырь. Оставаясь отверстыми, эти части вызывают образование уродств вроде Spina bifida или недоразвитие головного мозга (анэнцефалия). Подобные, а равно и некоторые другие уклонения могут быть вызваны в куриных яйцах, в белок которых был впрыснут раствор поваренной соли или других солей. Иногда, наоборот, раствор действует, вызывая усиленное разрастание зачатка: так, при действии растворов поваренной соли на яйца амфибий (Гертвиг, 1896), сахарного раствора на куриные (Шимкевич, 1902) замечается чрезвычайное разрастание нервного зачатка, а равно зачатков органов чувств (слуховых пузырей, хрусталика; см. Экзонейрула). Характер аномалий зависит отчасти от самого раствора, а главное — от природы яйца и от того, в какой стадии находится яйцо. Яйца морских звезд и ежей относятся различно к одним и тем же солям лития, и чем дальше в своем развитии зашло яйцо, тем менее оно подчиняется действию растворов, и в известных стадиях, вероятно, различных для различных животных, подавляющее действие растворов на отдельные зачатки прекращается вовсе. Наоборот, яйцо, подвергавшееся действию раствора в начале своего развития, будучи перенесено потом в чистую воду, все-таки дает ненормальных особей (Herbst, 1895). Влияние растворов во время дробления, если оно не ведет к общей задержке и полной остановке дробления, сказывается, главным образом, впоследствии. Впрочем, некоторые изменения могут происходить и во время самого дробления. Так, растворы некоторых солей, отнимая из различных частей яйца различное количество воды, тем самым могут изменять плотность различных частей яйца и через это влиять на форму дробления. Яйца миноги проходят нормально полное и неравномерное дробление, т. е. за двумя вертикальными бороздами следует 3-я горизонтальная, а в растворе некоторых солей 3-я борозда является тоже вертикальной (Bataillon, 1900). Надо, однако, отметить, что химический состав растворов играет весьма второстепенную роль и совершенно различные по химическому составу растворы оказывают часто сходное влияние. По-видимому, главная роль принадлежит здесь физическим свойствам того или другого раствора, а именно его способности проникать в клетки и вообще его осмотическим свойствам. Гербстом замечено, что чем более молекулярный вес той или другой соли данного металла, тем слабее ее влияние на развитие ее раствора. Чтобы получить тот же эффект, какой получается при действии 2,5 % раствора хлористого лития с литием бромистым, надо взять последний в 4 % растворе. Тому же закону подчинены и осмотические явления. Обратимся теперь к экспериментам над более поздними стадиями развития. Мы видели, что в стадии бластулы в полости ее скопляется жидкость. На яйцах пресноводного моллюска Physa Мезенгеймер (1896) показал, что при перенесении их в раствор морской соли сегментационная полость уменьшается и даже совсем исчезает. Точно так же и пресноводные простейшие, амебы и солнечники, при перенесении в соленую воду теряют свойственные им сократимые вакуоли. Очевидно, что пребывание в пресной воде вообще вызывает тенденцию к накоплению жидких продуктов в теле клетки или яйца, а в соленой воде эти продукты не накопляются, или, вернее, диффундируют прямо наружу. Замечательное изменение гаструляции амфибий в литиевых солях получил Гурвич (1896). Эктодерма, представленная у амфибии мелкими клетками верхнего полюса, останавливается в росте и не обрастает постепенно крупных, богатых желтком клеток нижней части яйца, как это бывает нормально. Вместо того, чтобы образоваться впячиванию на заднем конце яйца, образуется кольцевая борозда по всей окружности яйца. Механизм этого изменения не совсем выяснен. Аналогичное изменение, т. е. остановку роста эктодермического кружка (фиг. 6), лежащего на богатых желтком яйцах головоногих, удалось получить при развитии их в различных растворах, но там причина такой остановки иная и, по-видимому, чисто механическая: эктодермический кружок погружается своими краями прямо в желток, и последний препятствует его разрастанию, а это уже влияет на всю форму гаструляции (Шимкевич, 1900).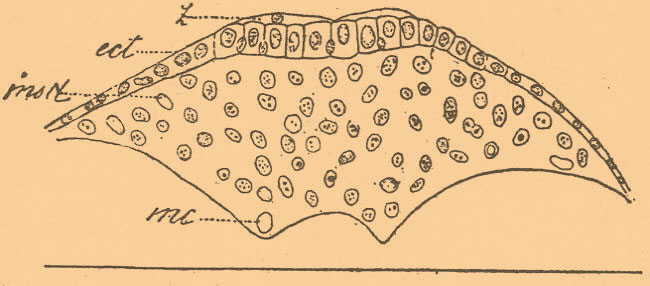
Фиг. 6. Раковинная железа зародыша Loligo vulgaris, развившегося в концентрированном растворе гуанина в течение 6 дней. Выползшие на поверхность железы клетки (g) приняли плоскую форму; ect — эктодерма; msd — сильно развитая мезодерма; mc — ядро мероцитов; желток не изображен (по Шимкевичу).
Затем, литиевые растворы вызывают получение экзогаструл, т. е. гаструл с вывороченным наружу кишечником, имеющих громадное значение для понимания процесса гаструляции (см. Экзогаструла). Аналогичное выпячиванию наружу кишечника явление наблюдается при выворачивании наружу нервной трубки цыпленка при убавлении белка (см. Экзонейрула). Точно так же хлористый литий, вызывающий выпячивание кишечника, оказывает то же действие и при развитии других органов. Так, слуховые пузырьки головоногих нормально появляются в виде полого эктодермического углубления (фиг. 3, А), а на одном зародыше, развивавшемся в растворе хлористого лития, этот пузырек выпятился наружу и представлялся в виде плотно сидящего на стебельке бугорка (фиг. 3, В).
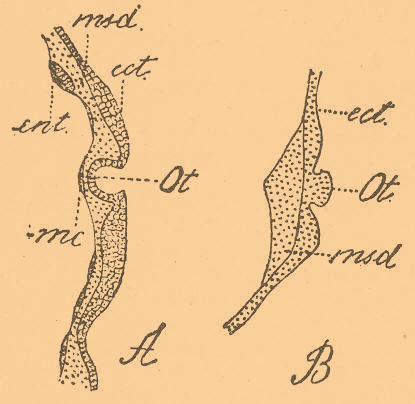
Фиг. 3. А — нормальное развитие статоциста у головоногих (по Бобрецкому); В — развитие статоциста у зародыша Loligo vulgaris, развившегося в течение 10 дней в 0,1 % растворе хлористого лития; еct — эктодерма, msd — мезодерма; ent — энтодерма; тс — мероциты; оt — статоцист; желток не изображен (по Шимкевичу).
Так, у всех моллюсков на спине эктодерма образует, так называемую, раковинную железу. У головоногих процесс этот совершается путем обычной инвагинации (фиг. 4).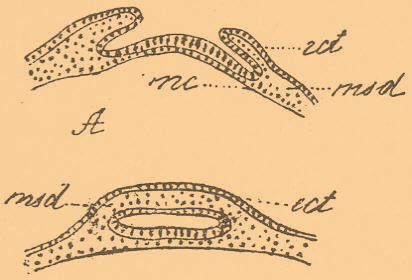
Фиг. 4. А и В — две стадии нормального развития раковинной железы у головоногих в разрезе; ect — эктодерма; msd — мезодерма; mc — слой мероцитов.
Но на зародышах, которые развивались в некоторых растворах и у которых эктодерма придавлена разросшейся мезодермой к оболочке, развитие раковинной железы происходит совершенно своеобразно. Клетки ее делаются цилиндрическими и обособляются по форме от соседних (фиг. 5), а затем по краям этого зачатка появляются вследствие размножения клеток эктодермы округлые клетки, которые выползают на поверхность зачатка, а затем принимают форму плоских кроющих клеток (фиг. 6) и наконец разрушаются.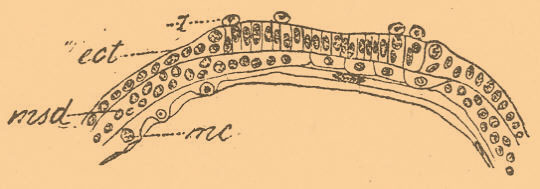
Фиг. 5. Раковинная железа зародыша Loligo vulgaris, развившегося в течение 3 дней в 0,5 % растворе йодистого калия; g — выползшие на поверхность ее клетки эктодермы — ect; msd — мезодерма; mc — мероциты, прилежащие к желтку, который не изображен (по Шимкевичу).
Процесс этот скорее должен быть сравнен с иммиграцией, соединенной с деламинацией, ибо является, в сущности, средним между ними. Другая интересная аномалия наблюдается на яйцах с экваториальной перетяжкой, образующейся довольно часто при развитии в растворах. Зародыш развивается только около верхней полусферы желтка, а нижняя остается обнаженной. Остановка разрастания зародышевого края вызывается своеобразным изменением роста, напоминающим телобластический способ развития. Если рассматривать край зародыша при сильном увеличении (фиг. 7), то оказывается, что ряды клеток эктодермы, упираясь в углубление на желтке, заканчиваются крупными клетками, находящимися часто в состоянии деления.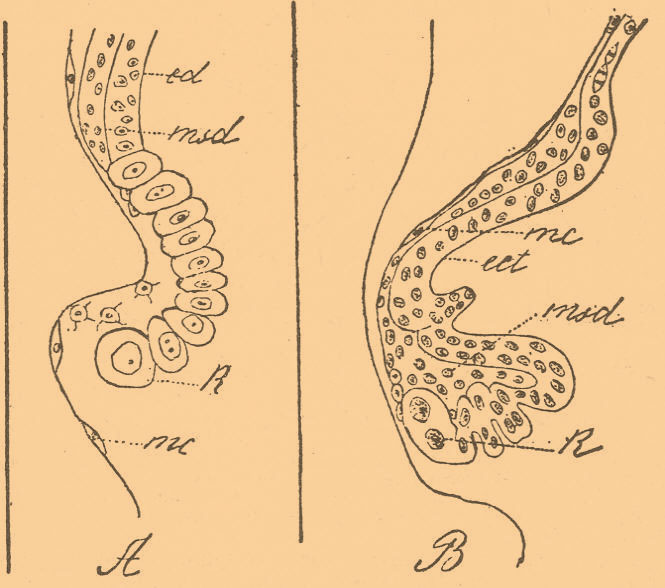
Фиг. 7. А — сильно увеличенный разрез через край зародышевого диска зародыша, развивавшегося в концентрированной морской воде в течение 8 дней; В — разрез через край такого же зародыша, но развивавшегося при тех же условиях в течение 5 дней; R — краевые клетки эктодермы — ect; msd — мезодерма; mc — мероциты, прилежащие к желтку, который не изображен (по Шимкевичу).
Но рост эктодермы идет лишь в одном направлении — кверху, где и образуется кругом яйца валик из эктодермы, иногда со складчатыми стенками. Глубже лежащие слои зародыша не представляют такого явления. Таким образом, клетки эктодермы, встречая препятствие и попадая в углубления желтка, где они находят условия усиленного питания, разрастаются и переходят к одностороннему телобластическому росту. Если вспомним, что, например, мезодермические телобласты у личинок лежат в задней части тела, где они, с одной стороны, встречают препятствие со стороны задней стенки зародыша и в то же время иногда и наилучшие условия питания, ибо всасывание пищи у личинок, вероятно, энергичнее происходит в задней части кишечника, то обнаружим некоторое сходство в условиях, в которых находятся телобласты личинки и краевые клетки описанного зародыша головоногого моллюска. Нетрудно также видеть, какое громадное значение все-таки имеет природа и состояние клетки: только клетки эктодермы, да и то не во всех стадиях, могут быть переведены к телобластическому росту, а прочие клетки, а именно мезодермические и прилежащий к желтку слой мероцитов, не представляют нам ничего подобного (Шимкевич, 1900). У цыпленка введением растворов в белок можно вызвать, кроме указанных изменений, целый ряд своеобразных уклонений: остановку роста зародышевого диска, подавление развития аллантоиса и амниона, а равно развитие последнего в отсутствие зародыша, изменение в расположении сосудов сосудистого поля, задержку замыкания бластопора, образование карликов, подавление развития конечности (см. Эктромелия), расщепление нервной трубки, задержку ее замыкания, экзонейрулы (см.), омфалоцефалию, или подгибание переднего конца нервной трубки к пупку, недоразвитие органов чувств, чрезвычайное развитие первичной полости, расширение полостей сомитов и целома, смещение (эктопию) сердца, водянку (гидропсию) сосудов, а равно и многие другие уклонения (Шимкевич, 1902). Интересны опыты Гербста (1903) над развитием яиц морских ежей в морской воде, лишенной той или другой из своих составных частей. Таким образом, по наблюдаемым уклонениям можно судить о том, какова роль той или другой составной части. Так, присутствие калия необходимо для роста зародыша, и в его отсутствие получаются маленькие бластулы, не образующие зачатка кишечника. Соли кальция необходимы для склеивания клеток зародыша, и в отсутствие этих солей бластомеры яйца расходятся, образуя неправильную кучку. Сульфаты также необходимы для роста, и их отсутствие вызывает гипертрофию ресничного пучка. Щелочной характер необходим для движения ресничек. В отсутствие хлоридов дробление начинается, но потом останавливается и т. п. Личинки морских ежей (Pluteus) снабжены отростками, в которых залегают известковые палочки скелета (фиг. 8, А).
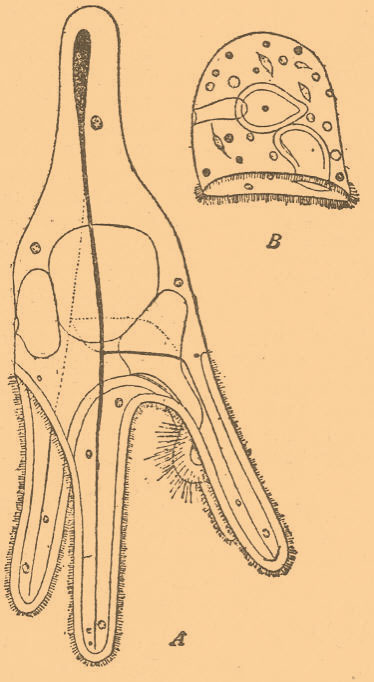
Фиг. 8. А — нормальная личинка (Pluteus) морского ежа в профиль (Strongylocentrotus); В — личинка другого ежа (Sphaerechinus), развившаяся в морской воде с примесью хлористого кали и лишенная скелета, в профиль; внутри той и другой личинки виден кишечный канал, состоящий из трех отделов: передней, средней и задней кишки (по Гербсту из Уильсона).
При отсутствии солей кальция в морской воде, а равно и вообще при ненормальном ее составе, вследствие примесей солей (Гербст), т. е. вследствие нарушения осмотических процессов, а равно при прибавлении к воде алкоголя (Циглер, 1903) отложения извести не происходит, а при этом не происходит и образования отростков (фиг. 8). Очевидно, что отростки возникают под влиянием раздражения, испытываемого тканями личинки со стороны известковых отложений. При развитии зародыша позвоночных возникающий кровеносный сосуд или нервное волокно, врастая в толщу мезодермы, вызывают около себя скопление клеток этого пласта, которые и облегают сосуд или нервное волокно, образуя их обкладку. Точно так же собираются около сосудов пигментные клетки. Леб (1896) показал, однако, что если прекратить ток крови в сосуде у зародыша рыбы (Fundulus), то блуждающие пигментные клетки не собираются около такого запустевшего сосуда. Очевидно, что привлекающим средством здесь влияет кровь или, вероятно, ее кислород. Точно так же возникновение хрусталика у позвоночных стимулируется прикосновением к эктодерме первичного глазного пузыря. Если развивается один такой пузырь, то и возникает один хрусталик, как это имеет место при некоторых случаях циклопии (см.). Отсюда возникло предположение о влиянии нервной системы на развитие органов. Однако это предположение оправдывается лишь отчасти. Леб перерезал в шейной части спинной мозг личинки аксолота, и метаморфоз задней части шел надлежащим порядком. Шапер (Schaper, 1898) утверждает, что у головастиков лягушки удаление головного мозга и органов чувств не влияет на ход превращения. Но на метаморфозе элементов, входящих в состав органов чувств, нервы, по-видимому, оказывают влияние. Вкусовые клетки, будучи изолированными от нервов, не атрофируются, а превращаются в обыкновенные эпителиальные (Semi Meyer, 1897). Также и при развитии органов чувств (Ехnеr, Hoffmann, Scymonowitsch) клетки эпителиальные превращаются в чувствующие, по-видимому, под влиянием подрастающих нервов. Таким образом, для органов чувств связь с нервами, по-видимому, является стимулирующей их развитие. Клейненберг (1886) еще давно, когда Э. эмбриология была в зачатке, указывал на некоторые случаи зависимости одного органа от другого, провизорного. Так, у личинок кольчатых червей появляется ряд провизорных органов чувств на головной лопасти. Все они потом исчезают, но каждый стимулирует появление под эктодермой кучки клеток, входящей в состав головного ганглия. Один орган замещает (субституирует) другой, вызывающий появление первого. Эти явления получили название явлений субституции, или подстановки органов. Итак, есть целая категория факторов, которыми мы можем объяснять некоторые явления развития, — это раздражения, вызываемые внешними (физико-химическими) факторами, и раздражения внутренние, вызываемые самими частями зародыша, действующими на другие его части. Производились многочисленные опыты удаления частей развивающегося яйца (см. Сегментационные шары). известны многочисленные случаи, когда изолированные бластомеры дают лишь часть зародыша, регенерирующую потом недостающие части (фиг. 9 и 10).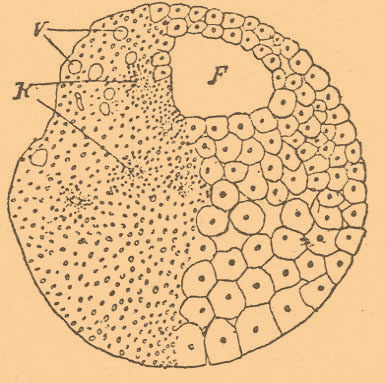
Фиг. 9. Полубластула, полученная после разрушения уколом одного из шаров дробления первой стадии лягушки, по Ру; F<
| "БРОКГАУЗ И ЕФРОН" >> "Э" >> "ЭК" >> "ЭКС" |
Статья про "Экспериментальная эмбриология" в словаре Брокгауза и Ефрона была прочитана 2282 раз
| Вкуснейшие куриные леденцы |
| Вкуснейшие куриные леденцы |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел