
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Череп, в анатомии
Определение "Череп, в анатомии" в словаре Брокгауза и Ефрона
Череп, в анатомии — костная или хрящевая коробка, в коей у позвоночных заключен головной мозг, некоторые органы чувств, а придатки этой коробки облекают передний отдел кишечного пути. В Ч. отличают, таким образом, спинную часть — собственно черепную или лучше краниальную, и брюшную часть внутренностную, или лучше висцеральную. Первая представляет облекающую головной мозг и органы чувств коробку, а вторая может быть рассматриваема как ряд дужек, частью видоизмененных, частью сохранивших форму дуг. Дужки эти называются висцеральными. Давно было замечено Океном и Гёте, что Ч. высших позвоночных может быть поделен на кольца, подобные позвонкам. Особенное сходство с позвонком обнаруживало заднее или затылочное кольцо. Отсюда возникла так называемая позвоночная теория Ч., по которой Ч. рассматривался как ряд видоизмененных позвонков, причем передние оказывались наиболее видоизмененными. Богатый костями Ч. костистых рыб, действительно, можно разложить на 4 кольца, как это и делал Оуэн, находя в этих кольцах части, соответствующие отдельным частям позвонка. Однако, число 4 оказалось слишком малым, если принять во внимание число черепных нервов (см.), и пришлось допустить, что в образовании Ч. участвовало большее число позвонков (Гегенбаур). Когда стали изучать развитие костей Ч., то оказалось, что они далеко не все однозначащие: одни развиваются из соединительной ткани и могут быть рассматриваемы как результат слияния базальных пластинок плакоидных чешуй (см.). Эти кости носят название накладных. Развитие, например, сошника и нёбных костей у амфибий, челюстных костей у двудышащих рыб наглядно показывает, что они возникли через слияние отдельных окостенений, несущих провизорные зубчики (у амфибий) или даже настоящие зубы (у двудышащих). Другие кости развиваются из надхрящницы первоначального хрящевого или первичного (примордиального) черепа и вытесняют собой этот последний. Кости эти называются эндохондральными. Сравнивать накладные кости с позвонком, коего все части развиваются эндохондрально, мы не имеем права. Если Ч. и представляет собой видоизмененную часть осевого скелета, то изменение это шло в ином направлении, чем изменение туловищной части: в последней развились позвонки, а в первой — черепные кости (Гексли). Изучение эмбрионального развития Ч. показало, что ни в одной стадии в нем не закладывается позвонков. Тем не менее, Ч. является таким же метамерным образованием, как и позвоночник. В туловище каждая метамера состоит из пары сомитов, пары нервов, со спинным и брюшным корешком каждый, и скелетной части (позвонка). В голове каждая метамера определяется точно так же парой сомитов, двумя парами нервов (а именно, парой спинных и парой брюшных ветвей) и затем соответствующей висцеральной дугой, тогда как краниальная часть остается не сегментированной. Но далеко не все эти элементы представлены в каждой метамере. Особенно много недостает брюшных нервных ветвей (см. Черепные нервы). Таким образом, позвоночная теория Ч. сменилась метамерной теорией. Для определения вопроса, сколько метамер входит в состав Ч., понадобилось точное исследование развития его у каждой группы, а это исследование привело к выводу, что число метамер, входящих в состав Ч. или лучше головы, у различных форм различно (Фрорип), но, говоря вообще, число это возрастает у высших позвоночных (Фюрбрингер, Северцов и др.). Мало того, возможно, что у одного и того же животного число мезодермических метамер головы не будет соответствовать числу метамер нервных или скелетных (Кольцов). В голове, так же, как и в туловище, мезодерма с каждой стороны делится на ряд сомитов и боковые пластинки, но в то время, как последние в туловище остаются не сегментированными, в голове они сегментируются под влиянием возникающих жаберных мешков (бранхиомерия), но эта сегментация, однако, не совпадает с метамерией, т. е. с делением на сомиты. Возможно, что это несовпадение есть явление вторичное. У круглоротых и селахий передние 2 пары сомитов возникают в виде полых выступов первичного кишечника, причем передний из них не имеет соответствующих боковых пластинок. Впереди слуховых пузырей у позвоночных образуются три пары сомитов — предушных (prootici), из них первый называется предчелюстным (praemandibularis), второй — челюстным (mandibularis) и ряд сомитов заушных (mettotici), лежащих позади слухового пузыря. Предушные сомиты, как и заушные, распадаются каждый на склеротом и миотом, причем миотомы их идут на образование двигающих глазами мышц. Строго говоря, у круглоротых в состав Ч. входят только склеротомы этих трех сомитов, но у селахий в образовании Ч. еще принимают участие склеротомы по крайней мере 6 пар заушных сомитов; таким образом, ван Вай (van Wijhe) насчитывает в голове селахий 9 метамер (а Дорн — гораздо больше); у осетровых число это возрастает еще больше (до 14) и еще больше у костистых (Северцов). У амфибий число головных сомитов невелико, но в этом случае возможно, что несколько затылочных сомитов исчезло без следа. В общем же, чем выше группа, тем больше число головных сомитов. Иногда можно обнаружить явственный след срастания Ч. с позвонками, соответствующими туловищным метамерам, а равно этот процесс отражается на увеличении числа черепных нервов (см.) вследствие присоединения к ним спинномозговых. Таким образом, около переднего конца хорды образуется из склеротомов скопление мезодермы, за счет коего и формируется Ч., коего часть, лежащая впереди хорды, называется прехордальной, а лежащая кругом нее — хордальной. Передний конец хорды оказывается включенным в заднюю часть нижней стенки Ч. Первая закладка Ч. обозначается закладкой в области его будущей нижней стенки 2-х пар хрящей, из коих передняя пара лежит впереди хорды (и только у круглоротых, двудышащих и амфибий они задними концами прикасаются к хорде), а задняя пара по бокам хорды. Передняя пара носит название trabecula cranii, а задняя — parachordalia, причем иногда ее зачаток является общим с хрящевыми капсулами, облекающими слуховые пузыри. Трабекулы могут лежать горизонтально или быть согнутыми под углом по отношению к parachordalia, что зависит от того, как изогнут головной мозг ко времени закладки трабекул. Затем все эти хрящи срастаются в одну пластинку, облекающую хорду и продырявленную отверстием между трабекулами, носящим название питуитарного. В этой области образуется потом гипофиз (gl. pituitaria), a впереди его сросшиеся трабекулы образуют пластинку, носящую название носорешетчатой перегородки (septum naso-ethmoidale). По бокам ее образуются капсулы, в которых помещаются органы обоняния. Этот хрящевой зачаток Ч., первоначально охватывающий нижнюю поверхность мозга, постепенно разрастается на бока его, а затем и на спинную поверхность, образуя капсулы для помещения органов обоняния, зрения и слуха. Питуитарное отверстие может закрываться еще в хрящевом Ч. или же закрывается позже накладной костью (os parasphenoideum). Все Ч. могут быть разделены на 2 типа: в одном случае правая и левая стенки межглазничной части Ч., обыкновенно вследствие сильного развития глазниц, сближаются между собой настолько, что эта часть уже не служит для помещения мозга, как бы отступившего кзади, и сближение этих стенок может быть настолько тесно, что здесь образуется тонкая перегородка, называемая межглазничной (septum interorbitale). Ч. этого типа имеют узкое основание и называются тропибазальными. Другой тип характеризуется широко расставленными стенками межглазничной части, заключающими в себе передний отдел головного мозга. Ч. этого типа имеют широкое основание и называются платибазальными. К числу последних относятся Ч. рыб и амфибий, а к числу первых — Ч. всех прочих позвоночных, не исключая и млекопитающих, у которых эта особенность несколько замаскирована (Гауп). Далее следует замещение хрящевого или первичного Ч. костным. Висцеральный скелет состоит из двух рядов хрящевых или позже окостеневающих дужек, из них передние две пары, мандибулярная и гиоидная, состоят из двух половин: верхней и нижней. Верхняя половина 1 дуги носит название небно-квадратного хряща (palatoquadratum) и играет роль верхней челюсти (см.) у низших рыб, а нижняя часть носит название мандибулярного или Меккелева хряща и тоже играет роль нижней челюсти или является пунктом, на котором формируются нижнечелюстные кости (см. Челюсти). Вторая дуга состоит из верхнего хряща (hyomandibulare) и нижнего (hyoideum), соединенных между собой непарной частью (basihyale). Впрочем, многие (ван Вай, Дорн) рассматривают эти два хряща, hyomandibulare и hyoideum, как самостоятельные дуги, некогда разделенные исчезнувшей ныне жаберной щелью. Между мандибулярной дугой лежит 1 жаберная щель (брызгальце рыб, среднее ухо (см.) прочих позвоночных). За этими дугами следует несколько пар дуг, лежащих позади соответствующих жаберных щелей и тоже соединенных непарными хрящиками на брюшной стороне (copula s. basibranchialia). Эти дуги называются жаберными или бранхиальными и число их бывает чаще 5, а у древнейших акул достигает 6 (Hexanchus и Chlamidoselache) и 7 (Heptanchus). Hyomandibulare уже у низших рыб играет роль подвеска (см. Челюстной подвесок), к которому причленяются челюсти, и у вышестоящих форм происходит ряд изменений висцерального аппарата. К изменениям Ч. по группам и переходим.
Ч. круглоротых рыб (табл., фиг. 2) отличается отсутствием челюстей, что стоит в связи с сосательным характером их рта, и является хрящевым или даже в верхней части перепончатым, причем срастается с непарной носовой капсулой, вытянутой, у Myxine, в длинную трубку, и слуховыми капсулами. По бокам Ч. имеются приросшие к нему рудименты дуг: нёбно-квадратной и гиоидной, а равно сильно развитая, подпирающая так называемый язык (см.), играющей роль поршня при сосании, непарная часть (basihyale). Бранхиальные дужки отодвинуты в туловище, что объясняется малым числом метамер, вошедших в состав Ч., и у Petromyzon соединены в цельную хрящевую решетку, заканчивающуюся капсулой, охватывающей сердце. Характерно также сильное развитие у круглоротых губных хрящей, поддерживающих ротовую воронку. Ч. селахий (табл., фиг. 3) тоже представляет хрящевую коробку, вытянутую спереди в выступ (rostrum) и тоже сверху впереди часто остающуюся перепончатой. Носовые капсулы приросли снизу к rostrum, а слуховые включены в толщу Ч., а между первыми и вторыми по выемке для глаз с отростками впереди глаз — анторбитальным и позади глаз — посторбитальным. С позвоночником Ч. соединяется или подвижно, или срастается с несколькими передними позвонками. Мандибулярная дуга представлена нёбно-квадратным хрящом, сросшимся с хрящом противоположной стороны и играющим роль верхней челюсти. С Ч. этот хрящ соединяется связками и только у Holocephala (см. Сростноголовые и Химеровые) срастается неподвижно с Ч. Гиоидная дуга представляет ряд градаций. У древнейших Notidanidae ее оба хряща составляют как бы одну дужку, несущую, подобно жаберным, хрящевые лучи, но у других акул нижний хрящ (hyoideum) — отступает назад, а у скатов (Rajidae) может лежать позади верхнего (hyomandibulare), который у Rajidae, будучи соединен связкой с мандибулярной дугой, является исключительно в виде челюстного подвеска (см.). У Holocephala hyomandibulare сливается с Ч. Гиоидная дужка распадается на ряд члеников: epihyale (сверху), keratohyale, hypohyale, а жаберные на pharyngo-, epi-, k e rato-, hypo-branchialia. Непарная часть задней дуги (copula) облекает снизу и с боков сердце (ср. капсулу круглоротых). Имеются рудименты губных хрящей. Хрящевой Ч. ганоидов (табл., фиг. 4) является замкнутым, но также спереди вытянут в rostrum, а сзади срастается с позвоночником. Но к этому Ч. сверху примыкает ряд накладных костей, у осетровых сохраняющих сходство с кожными отложениями, а равно на нижней поверхности Ч. развиваются две непарные накладные кости: сошник (vomer) и parasphenoideum. Точно так же имеются накладные, а равно эндохондральные кости в висцеральной части. Нёбно-квадратный хрящ играет роль верхней челюсти, но он уже одет костями. Мандибулярная дуга причленяется к hyomandibulare посредством промежуточного, обособившегося от последней хрящика — symplecticum. В жаберной крышке образуется окостенение (operculum). У других ганоидов накладные кости Ч. теряют свой кожный характер, и число их увеличивается. Ч. Amniota приближается к Ч. костистых рыб, а Ч. Polypterus — к Ч. амфибий. У костистых рыб (табл., фиг. 1) редукция хрящевого Ч. за немногими исключениями (Argyropelecus) весьма значительная, а равно развивается много костей как в краниальной части, так и в висцеральной, коих расположение дано на прилагаемой схеме. В висцеральной части небно-квадратный хрящ низведен на степень os quadratum, причленяющегося через os symplecticum к hyomandibulare и играющего тоже роль подвеска, а верхняя челюсть образуется парными накладными костями — praemaxillare и maxillare. Ряд костей развивается на мандибулярном хряще. Число костей в жаберной крышке, как и у многих ганоидов, значительно больше. Дужки также расчленены, а гиоидная несет еще ряд костяных лучей, подпирающих жаберную крышку (radii branchiostegi). Pharyngobrachialia жаберных дуг, будучи снабжены зубами, получают название верхнеглоточных костей, а задняя пара недоразвитых дуг — нижнеглоточных (ossa pharyngea superiora et inferiora). На прилагаемой схеме Ч. представлен сбоку и эндоходральные кости обозначены курсивом.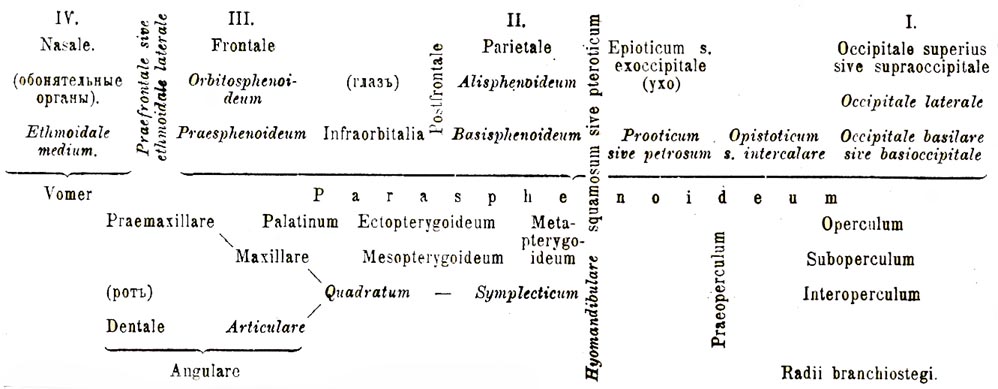
Ч. двудышащих (табл., фиг. 5) в значительной мере удерживает хрящевой характер. Характерно развитие некоторых костей, напоминающих амфибий, например лобно-теменных (fronto-parietalia), образованных каждая слиянием лобной и теменной. Нёбно-квадратный хрящ и рудиментарное hy o mandibulare слиты с Ч. и челюсть причленяется к косточке, считаемой одними за quadratum, другими — за squamosum. Жаберный аппарат и оперкулярный — сильно редуцированы. Сзади Ч. несет в затылочной части остистые отростки, что указывает на его слияние с несколькими (3) позвонками. Хрящевой Ч. амфибий (табл., фиг. 6) представляет коробку, открытую сверху, а иногда (Urodela) и снизу и дополняемую, поэтому, костями, не покрывающими, однако, всей поверхности хрящевого Ч. Висцеральный скелет характеризуется изменением верхнего участка гиоидной дуги (hyomandibulare и symplecticum) в слуховые косточки (columella и operculum, которое не надо смешивать с костью жаберной крышки; см. Слуховые косточки). На квадратном хряще, прирастающем к Ч., имеется отросток, который представляет различные степени развития. У Anura этот отросток, носящий название крыловидного (proc. pterygoideus), своим передним концом сливается с этмоидальной областью Ч. (ср. Petromyzon). Затем, в области барабанной перепонки имеется хрящевое кольцо (a n nulus tympanicus), на котором эта перепонка натянута и которое соответствует хрящу, наблюдаемому в области брызгальца у селахий. На этом хрящевом Ч. развивается ряд костей, по большей части встречаемых и у рыб. У Anura находим frontoparietalia, которые мы видели у Dipnoi, a равно в этмоидальной области особую кость (sphenoethmoideum), имеющую форму кольца и, может быть, образовавшуюся через смещение вперед и слияние обеих robitosphenoidea. У Gymnophiona (см. Червяги) и ископаемых Stegocephala число костей Ч. было больше. Своеобразные отношения представляет височная область, где образуется крыловидный отросток квадратного хряща. У амфибий на верхней стороне квадратного хряща развивается окостенение, которое одни считают за squamosum, другие — за особую кость (paraquadratum). Впереди от нее у Anura лежит косточка (quadrato-jugale s. quadrato-maxillare); которая упирается передним краем в максиллярную кость и образует скуловую дугу, прикрывающую височную область. У Urodela встречается такая же дуга, но она образована отростком paraquadrati и идущим навстречу ему отростком лобной кости. Первая дуга может быть названа нижней, а вторая верхней. У Tylotriton имеются обе дуги одновременно, а у Gymnophiona и Stegocephala кости височной области плотно прилежат к Ч. и дуг нет вовсе. Жаберный или подъязычный скелет Urodela, Gymnophiona и личинок Anura представляет непарную часть, образованную слиянием basihyalia и basibranchialia, и до 5-ти пар отростков, образованных гиоидной и жаберной дугами. У Anura редуцируются эти дуги, но зато вновь возникает несколько пар новых отростков, из коих передняя пара наибольшая. Боковые хрящи гортани Гегенбаур считает за производные 5-й пары жаберных дуг. У головастиков имеются губные хрящи, а равно их рудименты имеются у взрослых Anura. У рептилий хрящевой Ч. редуцируется в значительной степени, сохраняясь лишь в назоэтмоидальной области. Верхняя часть гиоидной дуги, как и у амфибий, представлена слуховыми косточками. Затем, у рептилий также имеется наклонность к образованию скуловых дуг. У морских черепах (см.) и некоторых ископаемых (Theromorpha) кости височной области плотно прилегают к Ч. и нет дуг. У Rhynchocephala (см. Хоботоголовые) и крокодилов имеются две дуги: верхняя и нижняя, но у большинства ящериц и отчасти черепах наблюдается одна верхняя дуга, а у змей, части ящериц и черепах — дуг нет вовсе. Дуги возникают вследствие редукции костного вещества между образующими дугу костями и стенкой самого Ч. Верхняя дуга образуется отростком squamosi и praefrontalia, а нижняя — jugale и лежащей позади нее косточкой, которая почти у всех рептилий соответствует, по-видимому, paraquadratum амфибий, и только у Hatteria она соответствует quadrato-jugale высших форм. В Ч. черепах бросается в глаза присутствие боковых полостей, образующихся вследствие разрастания на боках теменных костей и срастания их с ossa squamosa. Nasalia не развиваются, а носовая область, вследствие схождения внутренних (небных) отростков максиллярных костей с сошником, отделена от ротовой твердым нёбом (см.). Хоаны спереди ограничены сошником, а сзади нёбными костями. Quadratum вклинено в Ч. и сочленяется с нижней челюстью. У крокодилов (табл., фиг. 7 (1,3)), кроме наличности двух пар скуловых дуг, замечательно развитие твердого нёба: вследствие схождения отростков верхнечелюстных костей, нёбных и крыловидных костей, сошник закрыт, а хоаны отнесены далеко назад и окружены кругом крыловидными костями. Quadratum также вклинено в Ч., что замечается y Rhynchocephala. Между крыловидными и максиллярными костями имеется своеобразная кость — os transversum. У ящериц (табл., фиг. 7 (2,4)) бросаются в глаза боковые отростки в теменной области, существующие в сущности и у крокодилов, и от них отходит верхняя скуловая дуга. Теменные кости срослись и прободены foramen parietale (см. Теменное отверстие), встречаемым у Stegocephala и некоторых сомовых (Callichtys) и ископаемых ганоидов. Имеется os trasversum и еще столбчатая (у Rhynchocepliala в виде пластинки) кость, соединяющая крыловидные с теменными (epipterygoideum s. columella cranii). Ее нет у хамелеонов и амфисбен. Она представляет собой, вероятно, обособившийся отросток quadrati. Сошник парный и он, а равно нёбные и крыловидные кости, расходятся в бока, так что твердого нёба нет, a quadratum — сочленено подвижно с Ч. Отсутствие твердого неба и парность сошника характерна и для Ч. змей (табл., фиг. 8), у которых обыкновенно и sqamosum выклинено из Ч. и вошло в состав подвеска, вследствие чего достигается большая растяжимость челюстного аппарата, так как его отдельные кости соединены между собой растяжимыми связками. У неядовитых змей максиллярные кости удлинены, а у ядовитых цилиндричны и подвижно, наподобие шарнира, соединены с praefrontalia. Вследствие присутствия челюстно-нёбного аппарата (см.), образованного ossa transversa и крыловидно-небными костями, из коих первые упираются с одной стороны в quadratum, а вторые — в максиллярные кости, при раскрывании рта эти последние с их ядовитыми зубами выпрямляются, а при закрывании подгибаются. Подъязычный аппарат рептилий представлен непарной частью и двумя парами отростков (гиоидной и жаберной дужками). Он подвергается значительной редукции у змей.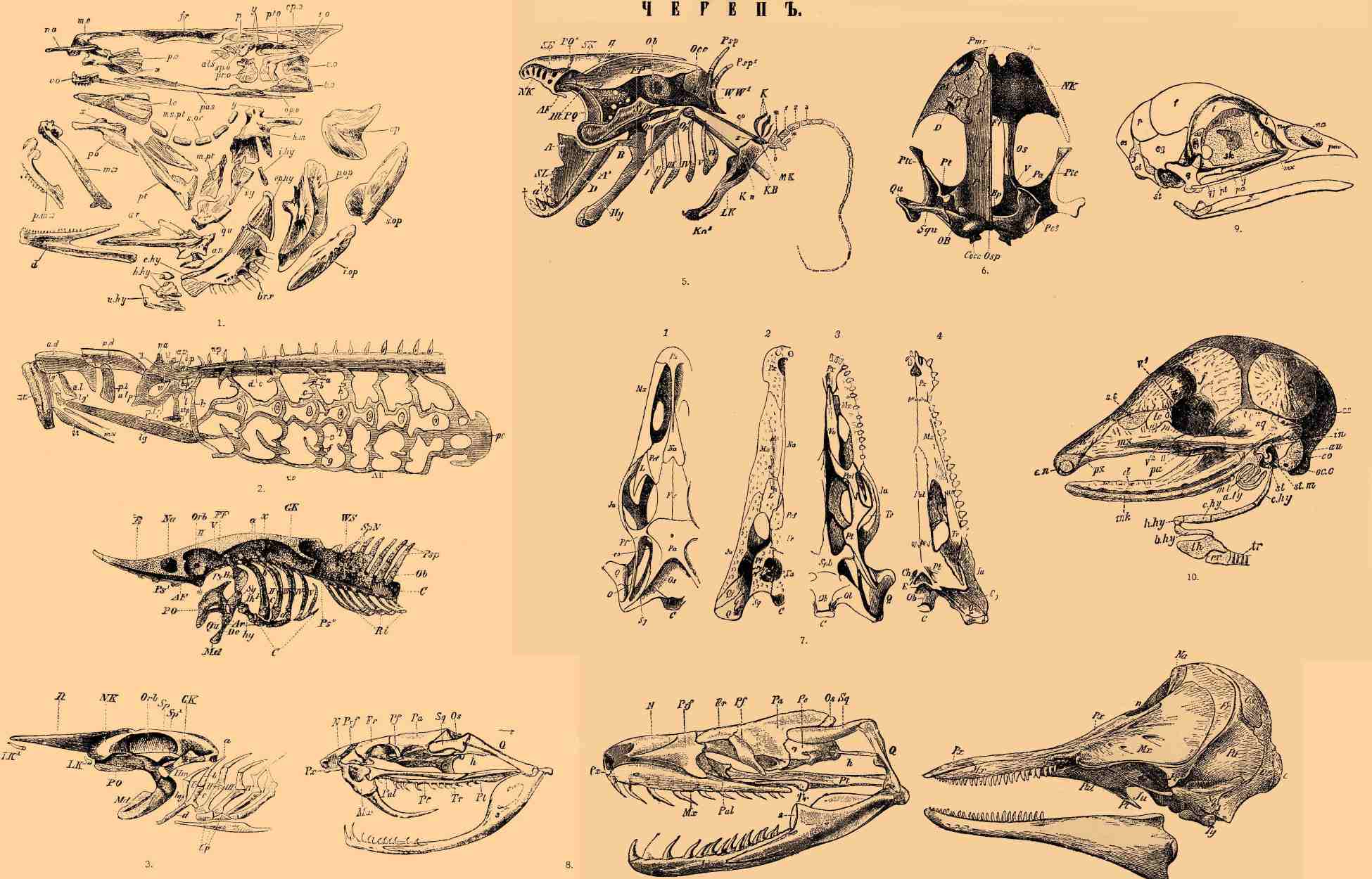
ЧЕРЕП.
К табл. ЧЕРЕП.
1. Череп трески (Gadus morrhua) с расставленными костями (из Паркера); als — alisphenoideum; an — angulare; ar - articulare; b.о — occipitale basilare; br.r — radii branchiostegi; c.hy — ceratohyale; d — dentale; e.o — occipitale laterale; ep.hy — epihyale; ep.o — epioticum; fr — frontale; h.hy — hypohyale; hm — hyomandibulare; i.hy — interhyale; i.op — interoperculare; lc — lacrymale; me — mesoethmoideum; m.pt и ms.pt — meta и meso-pterygoideum; mx — maxillare; na — nasale; op — operculare; op.o — opistooticum; p — parietale; pa.s — parasphenoideum; p.e — praefrontale; pa — palatina; p.mx — praemaxillare; p.op — praeoperculare; pr.o — prooticum; pt — ectopterygoideum; pt.o — squamosum; qu — quadratum; so — occipitale supenus; s.op — suboperculare; s.or — suborbitalia; sp.o — postfrontale; sy — symplecticum; u.hy — urohyale; vo — vomer; x, x', y и y' — сочленовные поверхности.
2. Череп морской миноги с передним отделом позвоночника и жаберным скелетом; nc — спинная струна с ее оболочками; np — остистые отростки; au — слуховая, na — носовая капсулы; bp, bp' — парахордальный, w — трабекулярный отделы черепа; II, V — отверстия для выхода зрительного и тройничного нервов; an — кольцевидный хрящ ротового отверстия; ad, pd, pl — добавочные подпорки последнего; zt, mw — губные хрящи; ztp, с.с, lg, lg', al — элементы гиоидной дуги; alp, plp — элементы pala to-quadrati; 1-7 — наружные жаберные отверстия; a-k — части жаберного скелета, d.c, v.с — его спинной и брюшной хрящи, параллельные спинной струне; pc — сердечная капсула (из Паркера).
3. Череп ската; GK — слуховая капсула; Orb — глазничная впадина; NK — обонятельная капсула; R — rostrum; LK, LK' — губные хрящи; Sp — хрящ брызгальца (Sp'); PQ — palato-quadratum; Md — нижняя челюсть; Hm — hyomandibulare; hy — hyoideum; I-V — жаберные дуги; а, b, с, d — их членики (pharyngo-, epi-, kerato— и hypobranchiale); † — место срастания последней и предпоследней жаберных дуг; Cp — copulae (из Видерсгейма).
4. Череп осетра; накладные кости удалены. WS — позвоночник; SpN — отверстия для выхода спинномозговых нервов; Psp — остистые отростки; Ob — верхние дуги; C — спинная струна; GK — слуховая капсула; AF, PF — пред— и заднеглазничный отросток; Orb — глазничная впадина; II — отверстие для зрительного, x — для блуждающего нервов; Na — носовая полость, R — rostrum; Ps, Ps1, Ps2 — parasphenoid; PQ — palato-quadratum; Q — quadratum; Md — нижняя челюсть; De — dentale; Ar — articulare; Hm — hyomandibulare; Sg — symplecticum; Ih — interhyale; hy — hyoid; I-V — жаберные дуги; a, b, c, d — их членики: pharyngo-, epi-, kerato— и hypobranchiale; Cop — copulae; Ri — ребра (из Видерсгейма).
5. Череп, плечевой пояс и передняя конечность Protopterus. W, W1 — входящие в состав черепа позвонки с их остистыми отростками (Psp, Psp1); Occ — supraoccipitale с отверстиями для прохода подъязычного нерва; Ob — слуховая капсула; Tr — trabeculae с отверстиями для тройничного и личного нервов; EP — fronto-parietale; HT — окошечко, прикрытое кожистой перепонкой с отверстием для прохода зрительного нерва (II); SK — окостеневшее сухожилие; SF — mesoethmoideum; NK — хрящевая носовая капсула; AF — предглазничный отросток (губной хрящ, имеющий такое же положение, не изображен); PQ — palatinum; Sq — squamosum (по другим quadratum); А, А 1 — articulare, соединенное с hyoideum (Hy) при посредстве фиброзной связки (B); D - dentale externum; †† — свободный отдел Меккелева хряща; SL — эмалевая пластинка; a.b — два зуба; OP, Op1 — рудиментарные кости оперкулярного аппарата; I-VI — шесть жаберных дуг; KB — черепное ребро; LK, MK — боковая и срединная костяные пластинки на хряще плечевого пояса (Kn, Kn1); co — фиброзная связка, прикрепляющая последний к черепу; x — сочленовный бугорок плечевого пояса; e — основной членик свободной конечности; 1, 2, 3 — три дальнейших членика последней; ** — рудимент ее боковых лучей; K — наружная жабра (из Видерсгейма).
6. Череп черной саламандры сверху. Cocc — затылочные мыщелки; Osp — спинной участок затылочного хряща; OB — слуховые пузырьки; Bp — хрящевая пластинка между ними; Squ — squamosum или, по другим, paraquadratum; Pot — processus oticus; Qu — quadratum; Ptc — хрящевое, Pt — костное pterygoideum; P и F — теменная и лобная кости; Pf — praefrontale, при D продырявленное слезно-носовым каналом; N — nasale; M — maxillare; Pmx — praemaxillare; Z — хрящевой вырост межносовой пластинки, служащей крышкой для cavum internasale; Na — наружное носовое отверстие; NK — обонятельная капсула; Fl — дыра для прохода обонятельного, V — тройничного нервов; Os — orbitosphenoideum (из Видерсгейма).
7. Череп ящерицы (1 и 8) и крокодила (2 и 4) сверху (1 и 2) и снизу (3 и 4). C — затылочный мыщелок; Ch — choanae; co — epipterygoideum; E — отверстие Евстахиевой трубы; Fr — лобная кость; Ju — jugale; L — слезная; Mx — верхнечелюстная; Na — носовая; Ob, Ol, Os — basi-, latero-, supraoccipitale; Pa — теменная кость; Pal — небная; Pf — postfrontale; Prf — praefrontale; Pt — крыловидная кость; Px — предчелюстная; Q — квадратная; Q', Qj — paraquadratum; Spb — basisphenoid; Sq — squamosum; Tr — transversum; Vo — сошник (из Гегенбаура).
8 (справа). Череп удава. Fr — frontale; h — stapes; Mx — maxillare; N — nasale; Os — supraeoccipitale; Pa — parietale; Pal — palatinum; Pe — petrosum; Pf — postfrontale; Prf — praefrontale; Pt — pterygoideum; Pk — praemaxillare; Q — qudratum; Sq — squamosum; Ts — transversum; 1, 2, 3 — кости нижней челюсти.
8 (слева). Череп ямкоголовой змеи (Craspedocephalus atrox). Обозначения те же, что и на предыдущем рис. Оба рис. из Боаса.
9. Череп двухдневного цыпленка сбоку. As — alisphenoideum; sk — межглазничная перегородка; x — кожистая часть ее; e — ethmoideum; na — хрящевые стенки обонятельной полости; pm — praemaxillare; f — frontale; p — раrietale; pa — palatinum; d — dentale; st — stapes sive columella; os — occipitale superius; ol — occipitale laterale; sq — squamosum; q — quadratum; qj — quadrato-jugale; pt — pterygoideum; pa — palatinum; mx — maxillare; n — nasale; l — lacrimale (из Боаса).
10. Череп Dasypus hybridus; хрящ изображен пунктиром (из Видерсгейма, по Паркеру). II — отверстие для выхода зрительного нерва; V1 и V2 — первая и вторая ветви тройничного нерва; a.my — annulus tympanicus; b.hy — basihyale; h.hy — hypohyale; c.hy — keratohyale; e.hy — epihyale; tr — trachea; cr — перстневидный хрящ; th — щитовидный хрящ; mk — Меккелев хрящ; d — dentale; ml — молоточек, in — наковальня; sx.m — stapes; an — слуховая капсула; oc.c - затылочный мыщелок; eo — occipitale laterale; s.о — supraoccipitale; sq — squamosum; p — parietale; i — jugale; pa — palatinum; f — frontale; lc — lacrymale; mx — maxillare; n — nasale; px — praemaxillare; en — наружное носовое отверстие.
11. Череп дельфина. C — condylus occipitalis; Oc — occipitale laterale; Os — supraoccipitale; Pa — parietale; Fr — frontale; Na — nasale; Mx — maxillare; Px — praemaxillare; Pal — palatinum; Pt — pterygoideum; Ju — jugale; Sq — squamosum; Ty — tympanicum; n — ноздри.
Для птиц характерно слияние многих костей Ч. вместе; quadratum сочленена подвижно с Ч., а равно имеется нижняя скуловая дуга. Эта дуга состоит из quadrato-jugale и jugale и упирается передним концом в верхнечелюстной аппарат, соединенный подвижно с этмоидальной областью, вследствие чего движения quadrati, имеющие место при раскрывании рта, сопровождаются (особенно у попугаев) поднятием верхнечелюстного аппарата. Кроме того, ossa quadrata соединяются с верхнечелюстным аппаратом посредством крыловидно-нёбных костей, упирающихся в то же время в выступ базальной части Ч. или rostrum и скользящих по нему. Как скуловая дуга, так и нёбно-крыловидный рычаг являются передатчиками движений quadrati верхнечелюстному аппарату и одевающему его надклювью. Подъязычный аппарат с сильно развитой непарной частью и двумя парами отростков, как у рептилий. Характерным является для Ч. млекопитающих превращение quadrati и articulare в слуховые косточки (наковальню и молоточек). Затем, в то время как сочленение Ч. с позвоночником у амфибий имеет тот же характер, как и сочленение позвонков между собой, а у рептилий и птиц оно совершается при помощи непарного мыщелка, образованного 3 костями: occipitale basilare и 2 occipitalia lateralia, y позвоночных сочленение это совершается при помощи 2 мыщелков, образованных этими последними костями. Однако, нетрудно себе представить происхождение этого непарного мыщелка из непарного мыщелка рептилий, если допустить, что occipitale basilare перестало принимать участие в образовании его срединной части. Все кости черепа млекопитающих соединены, кроме слуховых, подъязычной и нижней челюсти, неподвижно, а затем те кости, которые являются изолированными в Ч. нижестоящих форм, сливаются вместе и образуют сложные кости, а в редких случаях (Monotremata, Chi r optera) кости Ч. вообще сливаются между собой. Рассматривая с этой точки зрения человеческий Ч., мы найдем следующее: сзади Ч. ограничен костью (os occipitale), представляющей продукт слияния occipitale basilare, lateralia и superius. У сумчатых эти кости могут быть отделены одна от другой. Occipitalia lateralia несут мыщелки для сочленения с атлантом и у многих форм снабжены особым отростком каждое — processus paramastoideus. Точно так же у приматов и хищников с затылочной костью сливается особая, соединительно-тканная, парная по происхождению кость, os interparietale; иногда (Cetacea) затылочная кость, вследствие этого слияния, доходит своим передним краем до лобной, отделяя обе теменные кости друг от друга. В других случаях interparietalia остаются свободными или сливаются с теменными (грызуны и жвачные). Сбоку Ч. млекопитающих ограничен височной костью — os temporale, тоже весьма сложного характера, а именно: ее pars petrosa соответствует os petrosum, pars squamosa — os squamosum, которая, впрочем, может оставаться самостоятельной, а стенки слухового прохода образованы os tympanicum. Каменистая кость млекопитающих, petrosum, происходит за счет сливающихся друг с другом pro-, epi— и opisthoticum и заключает в своей толще слуховой лабиринт. Она принимает значительное участие в образовании черепной кости, а именно ее дна, но на наружную поверхность Ч. почти не выходит. Происходит это потому, что снаружи с ней срастается имеющая собственный центр окостенения сосцевидная кость — os masioideum, несущая у человека processus mastoideus, и еще другая, подобно squamosum, тоже соединительно-тканного происхождения, барабанная кость tympanicum. Только перед передним краем этой последней petrosum узкой полоской выступает наружу, причем именно в этом месте до полного срастания барабанной и каменистой кости дело не доходит и образуется так называемая Глазерова щель (fissura Glaseri); tympanicum, по Гегенбауру, соответствующая quadrato-jugale, в простейшем случае (Monotremata, Marsupialia, некоторые Insectivora) является в виде неполного кольца, на котором натянута барабанная перепонка. В других случаях кольцо это, разрастаясь, вытягивается в более или менее длинную трубку, образующую нередко (Rodentia, Carnivora, Perissodactyla) при основании Ч. особый пузырь (bulla tympanic a), играющий роль резонатора. Сходное образование имеется и у Marsupialia, но там оно развивается за счет alisphenoidea. Squamosum характеризуется особым направленным вперед отростком — processus zygomaticus; этот последний у насекомоядных Sorex и Centetes кончается свободно; у неполнозубых Myrmecophaga и Bradypus связан со скуловой костью (jugale) посредством особой связки, а у всех прочих Mammalia вступает со скуловой костью, jugale s. zygomaticum, a через последнюю и с верхней челюстью в непосредственную связь, и таким образом формируется верхняя скуловая дуга. Далее, кпереди, в основании Ч. мы находим у человека основную кость (os sphenoideum). Она представляет собой результат слияния, во-первых, basisphenoidei и alisphenoidea, причем последние образуют большие крылья (alae magnae), a во-вторых, praesphenoidei и orbithosphenoidea, образующих alae parvae. Наконец, внутренняя пластинка processus pterygoidei образована из ossa pterygoidea, у большинства остающихся самостоятельными. У большинства, однако, мы находим, что basisphenoideum со своими крыльями остается свободным от передней части основной кости, или praesphenoideum, снабженного alae parvae, иногда превышающими величиной alae magnae. Передняя часть дна черепной коробки образована решетчатой (ethmoid e um) костью, развивающейся из двух боковых и одного непарного окостенения, причем последнее соответствует lamina perpendicularis. Lamina perpendicularis вдается в Ч. в виде crista galli, a боковые части обращены в черепную полость решетчатыми пластинками, имеющими каждая у утконоса одно, а у прочих массу мелких отверстий для прохождения nervi olfactorii; боковые части этой кости образуют ячеистые массы или лабиринты. От каждого лабиринта свешивается в обонятельную полость по два пластинчатых выступа, служащих, как и самые лабиринты, для увеличения обонятельной поверхности и весьма варьирующих у различных Mammalia с точки зрения формы и величины. Эти так называемые этмоидальные раковины (Ethmoturbinalia) — образования, свойственные млекопитающим исключительно. Напротив того, что до нижней носовой раковины, os maxillo-turbinale, свойственной рептилиям и птицам, то она окостеневает самостоятельно, но часто срастается с максиллярными костями. Parietalia иногда срастается с frontalia, тоже могущими часто сливаться между собой. Явление несрастания frontalia у человека носит название метопизма, и чаще встречается у высших рас, чем у низших. Кроме этих костей, мы находим еще в лицевой части Ч. непарный, но слившийся из двух половин, сошник, который в отличие от рептилий лежит не горизонтально, а вертикально, образуя часть носовой перегородки; 2 скуловые кости, иногда (Centetes, Sorex) отсутствующие, а также еще 2 отсутствующие у ластоногих и дельфинов слезные кости — lacrymalia. Слезные кости нередко сильно развиты и выходят на наружную лицевую поверхность, будучи снабжены у многих жвачных углублениями для помещения особых сальных желёз. Носовая полость является снизу ограниченной сошником; с боков — челюстными костями, сверху прикрыта двумя соединительно-тканными носовыми костями (nasalia), отсутствующими у Sirenia, рудиментарными вследствие перемещения ноздрей кзади у Cetacea и слонов, иногда же срастающимися вместе (носороги, обезьяны Старого Света). Носовая перегородка образована lamina perpendicularis решетчатой кости и сошником. В области носа остаются участки хрящевого Ч., образующие хрящи в носовой перегородке и крыльях носа. У Primates, Perissodactyla, жвачных и бегемота скуловая кость дает отросток, соединяющийся с особым отростком лобной кости, и отграничивает полость орбиты от височной впадины Ч. костяной дугой. У Primates дело идет еще далее: отверстие между этой дугой и стенкой Ч., именно alae magnae, замыкается вследствие разрастания последних, так что височная ямка и орбита соединяются уже не широким отверстием, а узкой щелью (fissura orbitalis inferior). Полость глазницы у Primates является образованной, кроме лобной кости, большими крыльями, скуловой костью, отростком верхней челюсти, затем слезными, всецело лежащими в глазнице костями и боковыми частями e t hmoidei, образующими так называемую lamina papyracea, особенность, свойственная еще некоторым Edentata. У других же форм, где не происходит соединения скуловой кости с лобной, глазница от височной ямки не отделена вовсе. Сверху и спереди ротовая щель ограничена praemaxillaria, которые, за исключением антропоморфных и человека, являются самостоятельными костями и легко могут быть найдены на Ч. Прекрасно развитые у Rodentia и Proboscidea (особенно у первых), в других случаях (некоторые Edentata и Chiroptera) они являются слаборазвитыми. Следующие кзади от praemaxillaria maxillaria у низших Mammalia сильно развиты и вытянуты вперед, у высших же, так сказать, подтянуты кзади. У Cetacea они настолько вытянуты назад, что закрывают собой лобные кости. Их небные пластинчатые отростки сближены по срединной линии и вместе с palatina образуют твердое нёбо — palatum durum. Ossa pterygoidea, у человека и обезьян срастающиеся с основной костью, только у Cetacea и Edentata принимают участие в образовании твердого нёба, причем у муравьедов, броненосцев и некоторых китов хоаны отодвинуты назад, как у крокодилов. Нижняя челюсть соответствует только сросшимся вместе ossa dentalia прочих позвоночных и сочленяется с os squamosum. Наверху имеет венечный отросток (pr. coronoideus), иногда отсутствующий (Cetacea и др.). Гиоидная дужка сохраняется частью в виде передних рожков подъязычной кости, а верхний ее участок у человека и орангутанга, окостеневая и прирастая к petrosum, образует pr. styloideus, или шиловидный отросток, соединенный связкой с передним рожком. Первая жаберная дуга образует задние рожки подъязычной кости, прикрепленные к гортанным хрящам, а прочие части висцерального аппарата сохраняются в виде гортанных хрящей (см. Черпаловидные хрящи и Щитовидный хрящ). У Monotrem a ta подъязычный аппарат и щитовидный хрящ находятся в тесной связи. Кости человеческого Ч. описаны под соответствующими названиями.
Литература. Кроме более старых работ: Owen, 1855; Huxley, 1864; Gegenbaur, 1887, важное значение и
| "БРОКГАУЗ И ЕФРОН" >> "Ч" >> "ЧЕ" >> "ЧЕР" >> "ЧЕРЕ" |
Статья про "Череп, в анатомии" в словаре Брокгауза и Ефрона была прочитана 1294 раз
| Коптим скумбрию в коробке |
| Панайпай |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел