
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Человек, зоологический очерк
Определение "Человек, зоологический очерк" в словаре Брокгауза и Ефрона
Человек, зоологический очерк
Человек, зоологический очерк
(Homo sapiens) — Так как отдельные органы и части человеческого тела описаны под соответствующими названиями, то в настоящей статье мы будем говорить о Ч. только с точки зрения зоологической, т. е. относительно его отличия от ближайших к нему животных. Что касается взглядов, существующих на счет места занимаемого Ч . в системе, то они довольно разнообразны. Некоторые французские антропологи устанавливают для Ч. особое царство, характеризующееся явлениями религии, морали и членораздельной речью, а другие — не находят возможным отделять род Homo от семейства человекообразных обезьян (см.). Между этими крайними взглядами укладываются все прочие, как-то: выделение Ч. в особое семейство Homin i dae, затем в особый подотряд приматов или возведение на степень самостоятельного отряда (Блуменбаховский отряд Bimana или двуруких), параллельного приматам, или даже на степень самостоятельного класса (К. Г. Карус). Линней в своей "Systema naturae", в позднейших изданиях, определял положение Ч. в системе следующим образом:
2. Simia.
3. Lemur.
4. Vespertilio.
Иначе говоря, Линней соединял с приматами летучих мышей (Vespertilio) и полуобезьян (Lemur), теперь отделяемых в особые отряды, но зато антропоморфных обезьян (H. sylvestris) он ставил ближе к Ч., чем к прочим обезьянам (Simia). Под именем H. monstruosus он соединял большинство уродливостей и аномалий, а под именем H. ferus разумелись волосатые люди и т. п. аномалии. Прочие деления Линнея представляют попытку разделения человеческого рода на расы (см.). Ламарк уже рисует нам гипотетическую картину того, как из "расы четвероруких" возник Ч. (см. Трансформизм). Появление книги Дарвина о происхождении человека явилось следствием общего взгляда на эволюцию всего организованного мира и поставило вопрос ребром. Гексли уже высказывает положение, что Ч. по своей организации гораздо менее разнится от высших обезьян, чем эти последние от низших, и что, если бы обитатель Сатурна спустился на землю, то он не задумался бы относительно места Ч. в системе животных, населяющих землю. Развитию и распространению взглядов Дарвина много содействовали Геккель и К. Фогт. Как бы то ни было, при решении вопроса о положении Ч. в системе, надо иметь в виду только его анатомическое строение, и с этой точки зрения даже блуменбаховский отряд Bimana оказался не выдерживающим критики, ибо задняя конечность обезьян представляет собой не руку, а типичную ногу, и они являются такими же двурукими, как и Ч. Что же касается до умственных отличий Ч. от. животных, то они, как это формулировано Дарвиным, являются количественными, а не качественными. Немало отнял сил и времени с современной точки зрения довольно бесплодный спор между моногенистами и полигенистами о том, представляют ли все ныне живущие человеческие расы один зоологический вид, как это утверждают первые, или два и более видов, как это утверждают вторые, а, сообразно этому, решался и вопрос о возникновении Ч. из одного источника или из нескольких. Мы знаем теперь, что понятие о виде и разновидности настолько условно, что спор этот, будучи поставлен на чисто таксономическую почву, почти не имеет интереса. Действительно, расы, населяющие Южную Африку, Австралию и часть Полинезии, могут быть противопоставлены расам, населяющим С. Африку, Европу, Азию и обе Америки, но разница между этими двумя группами все-таки не более разницы между двумя разновидностями. Возможно, что эти разновидности были обособлены уже в делювиальную эпоху, но все известные нам ископаемые остатки, по-видимому, принадлежат предкам второй разновидности, а делювиальные предки первой разновидности неизвестны. Таким образом, все ныне живущие и большая часть ископаемых рас соединяются под именем Н. sapiens, но одна ископаемая раса отделяется в особый вид H. nean derthalensis.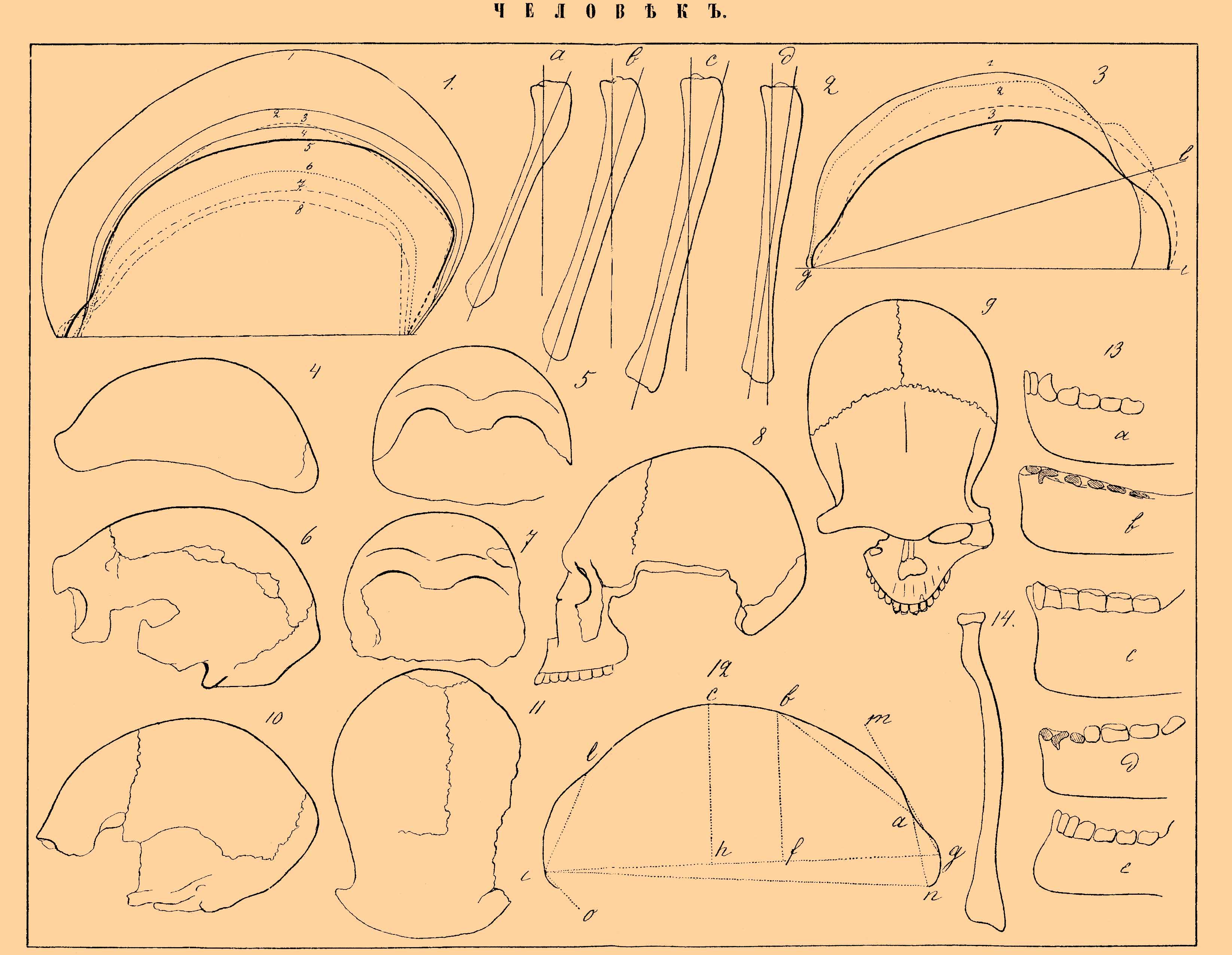
ЧЕЛОВЕК (Зоология)
Фиг. 1. Профили черепов (в проекции): 1 — папуаса; 2 — Спи № 2; 3 — (пунктиром) — микроцефала (случай Куннингама); 4 — Спи № l; 5 — Pithecanthropus; 6 — Гиббона (Hylobates leuciscus); 7 — Semnopithecus maurus; 8 — шимпанзе (Anthropithecus). Величина черепов уменьшена вдвое (по Дюбуа). Фиг. 2. Схематичные контуры больших берцовых костей (tibia) с внутренней стороны и положение их осей: а — взрослой самки гориллы; b — Спи № 1; с - каменного века (неолитического периода); d - современного жителя Льежа (по Фрэпону). Фиг. 3. Профили черепов (в проекции); 1 — современного эльзасца; 2 — эгисгеймского; 3 — неандертальского; 4 — Спи № 1 (по Клаачу); g — i — линия, соединяющая глабеллу с инион (glabella в смысле Брока — валик, лежащий над корнем носа в наиболее выдающейся его части; inion — в смысле Меркеля — затылочный гребень, т. е. protoberantia occipitalis externa); g — l — g, линия, соединяющая глабеллу с ламбдой (lambda — наиболее высокий пункт затылочно-теменного шва). Фиг. 4. Профиль неандертальского черепа. Фиг. 5. Он же спереди. Фиг. 6. Профиль черепа Спи № 1. Фиг. 7. Он же спереди. Фиг. 8. Череп Самбаки (около Сантос) в профиль. Фиг. 9. Он же сверху (по Нерингу). Фиг. 10. Профиль черепа Спи № 2. Фиг. 11. Череп Спи № 1 сверху. Фиг. 12. Профиль неандертальского черепа (в проекции): о — опистион (Opisthion — точка на заднем крае затылочного отверстия); i — ннион; l - ламбда; g — глабелла (в смысле точки, лежащей между надбровными дугами); n — назион (nasion — наиболее выдающаяся точка лобно-носового шва); b - брегма (bregma, место соединения лобной и двух теменных костей); а - fossa supraglalellaris (углубление над глабеллой); с — наивысшая точка сагиттальной кривой; bf — высота брегмы; mgi - лобный угол. Фиг. 13. Профили нижних челюстей: а — шимпанзе; b — ноллетской; с — Спи; d — жителя новогибридских островов; е — парижанина. Фиг. 14. Лучевая кость неандертальского скелета.
Мы приходим к необходимости познакомиться с ископаемыми остатками Ч. Весьма важным является вопрос о древности человеческого рода. Надо отметить, что по отношению к Ч. определение геологической древности тех напластований, в которых залегают остатки, должно быть принимаемо с большой осторожностью. Ч., вероятно, и в доисторические времена зарывал своих покойников, так что нахождение человеческих остатков в данном пласте еще не определяет вполне достоверно геологического возраста этих остатков, ибо они могли попасть в него при погребении. Точно так же одновременное нахождение костей Ч. в пещерах вместе с костями вымерших животных не всегда является доказательством одновременного существования этих остатков, ибо этими пещерами Ч. мог также пользоваться для погребения покойников в более позднее время. Поэтому относительно делювиального характера некоторых остатков, напр., черепов Spy, канштадтского и даже неандертальского имеются возражения, и некоторые исследователи считают их черепами позднейшего происхождения. Является вопрос, существовал ли Ч. в третичную эпоху. Мы не имеем ни одного достоверного остатка человеческого скелета, принадлежащего к третичной эпохе. Найденный в третичных отложениях Калифорнии череп (Calaveras-Schädel) принадлежит индейскому покойнику. Найденные в Пампасах Южной Америки человеческие остатки (резцы в нижних слоях, черепа и скелеты в верхних) принадлежат делювиальным наслоениям, а не третичным. Найденные в Северной Америке в третичных слоях около Карона отпечатки следов, принятые за человеческие, принадлежат, по-видимому, какому-то четвероногому, а найденные в Австралии около Виктории отпечатки следов и сидений Ч., хотя и представляют громадный интерес, ибо показывают, что в начале делювия Австралия была уже обитаема Ч., все-таки не могут быть отнесены с полной достоверностью к третичным слоям. В пользу древней обитаемости Австралии Ч. говорит и то обстоятельство, что в делювиальных ее отложениях найдены остатки скелетов динго, одичавшей и несомненно привезенной в Австралию человеком формы. Затем найдены иного рода следы пребывания Ч. в третичную эпоху. Так найдены были кости некоторых животных, носящие следы повреждений, как будто каменным орудием. Однако зубы крупных грызунов, удары пилы-рыбы и т. п. могут, по-видимому, также произвести подобные же повреждения. Были найдены кремни, носящие следы грубой обделки ударами или, по другим, треснувшие от нагревания на огне, но действительно ли эти кремни подвергались выделке — не ясно. Такие первобытные орудия найдены Нётлингом в Индии вместе с зубом третичного предка лошади (Hippotherium antelopinum), что заставляет отнести их появление, если не к миоцену, то к плиоцену. Если третичным предкам Ч. мы не можем приписать выделку настоящих кремневых орудий, то все же можно допустить, как это предполагает Рюто (Rutot), что они пользовались найденными ими осколками подходящей формы и, может быть, несколько приспособляли эти кремни обколачиванием. Во всяком случае в начале четвертичной эпохи Ч. уже существовал в форме, весьма близкой к современной. Попытки вычислить время существования Ч. на земле не могут опираться на сколько-нибудь прочные данные, и потому в то время, как одни высчитывают древность Ч. в 15000—20000 лет, другие считают 200000 лет. От делювиального Ч. мы имеем каменные орудия, расколотые и вообще поврежденные кости животных и, наконец, скелеты. По-видимому, Ч. тогда вел образ жизни дикаря и питался, главным образом, мясом убитых животных. В Северной Америке до сих пор не найдено остатков делювиального Ч., кроме куска бедерной кости и немногих каменных орудий, ибо большинство североамериканских орудий, принимавшихся за делювиальные, позднейшего происхождения и представляют собой отбросы при выделке каменных орудий индейцами. В Южной Америке, мы видели, найдено несколько черепов и скелетов, найдены следы делювиального Ч. в Австралии, но главные находки сделаны в Европе. Вообще же по сравнению с делювиальной фауной человеческих остатков найдено мало. Бранко объясняет это тем, что тогда вообще число представителей человеческого рода было невелико и заселение земли шло медленно, так как масса людей погибала от голода, болезней и войны. Надо думать, что возник человеческий род в тропиках и постепенно распространился по всему свету, заходя вслед за мамонтом и волосатым носорогом на отдаленный север. Наибольшую роль в суждении относительно происхождения Ч. играл так назыв. неандертальский череп, найденный Фульротом и Шафгаузеном в 1859 г. в Неандерской долине близ Дюссельдорфа. Найдена собственно черепная крышка и некоторые кости конечностей. Череп (фиг. 4 и 5) этот большой, длинный и в то же время широкий и кажущийся еще более длинным оттого, что его надбровные дуги сильно выдаются вперед; лоб отступающий, отклоняющийся сильно кзади, вследствие чего черепная крышка является низкой, а равно и мозговая полость была малой величины. Промежуток между полостями обоих глазниц был большой. Конечно, мнения относительно этого черепа разделились. Одни полагали, что это — череп недавнего происхождения, причем было обнаружено, что между современными черепами встречаются сходные с неандертальским. Так, в Юрьевском университете имеется череп, найденный при раскопках в Эстляндской губ. и сходный с неандертальскими. Конечно, наиболее естественным объяснением такого явления было бы признание в современном Ч. атавистической тенденции и возврата к неандертальскому типу, но много содействовал затемнению вопроса Вирхов, который, как патолог, всюду находил болезненные уклонения и применил эту точку зрения к неандертальскому черепу. В настоящее время, когда череп этот был снова переисследован Швальбе, басня о его патологическом характере падает. Этой басне содействовало, между прочим, то обстоятельство, что чрезвычайно распространенные по всем музеям слепки черепа были не совсем верны, а именно, они преувеличивали толщину черепной стенки, а когда с черепа была очищена земля, то толщина костей оказалась не такой значительной. Затем другие кости, найденные вместе с неандертальским черепом и исследованные недавно Клаачем, не представляют никакого следа патологических процессов. Когда потом были найдены черепа Ларте в 1860 г. в пещере Spy в Бельгии, из которых один особенно близок к неандертальскому, а потом и другие, тоже к нему отчасти приближающиеся, то пришлось или считать их все патологическими, или отказаться от этой точки зрения совсем. Кинг и Копе предложили выделить неандертальский тип в особый вид Ноmо neanderthalensis, с чем согласен и Швальбе, присоединяющий сюда же и черепа Spy. Исследование Швальбе черепов неандертальского и черепов Spy привело его к следующим результатам:
1) высота черепов Н. и Spy (коей показатель определяется согласно прилагаемой фиг. 12 (ch×100)/(gi) значительно ниже черепов наиболее низко стоящих рас, а именно высотный показатель первых = от 43,7 до 46,4, тогда как minimum современных черепов = 52, a maximum для шимпанзе 38; высотный показатель неандертальского черепа = 40,4, иначе говоря, по этому признаку Н. neanderthalensis стоял ближе к антропоморфным, чем к Ч., 2) брегматический угол bgi (фиг. 12) 44°, а у Homo sapiens minimum 53°, у шимпанзе 39,5°; 3) показатель положения брегмы (см. объяснение) (fg×100)/(ig) на неандертальском черепе = 38, у Н. sapiens maximum — 34, у оранга minimum — 43,5; 4) лобный угол (moi) на неандертальском черепе = 62°, у H. sapiens minimum = 80°, у шимпанзе maximum = 56°; эти цифры определяются, главным образом, формой лба, сильно отклоняющегося кзади у H. neanderthalensis; 5) по сильному выступу надглазничной части черепа в виде torus supraarbitalis также H. neand. приближается к обезьяньему, а именно линия an только в два раза короче линии bа, a у Н. sapiens в 4 — 5 раз короче; 7) длина теменной кости по срединной линии у H. neander. и антромофорных обезьян короче таковой же длины лобной кости, а у Н. sapiens — наоборот; 7) величина угла lig на неандертальском черепе = 66,5°, на черепах Spy 68°, что опять-таки сближает эти черепа с обезьяньими, где этот угол варьирует от 43° до 68°, тогда как у Н. sapiens он варьирует от 78° до 85°.
Сравнительное отношение размеров черепов неандертальского, Spy и современных эльзасцев выясняется следующими цифрами:
| Spy № 1 | Spy № 2 | Неандертальский | Современный | |
| Длина (в мм) Ширина Длина линии gi (glabellainion) Высота (ch) Высотный показатель Длина линии gl (glabella-lambda) | 200 140 196,5 82 41,7 182 | 198 150 190 87 45,7 185 | 202 144 196 91 40,4 192 | 155 — 201 125 — 165 — 91 — 106 55 — 64 — |
Долготно-широтный показатель (см. ниже) Spy № 1 = 70; неандертальский череп = 72; Spy № 2 = 74; следовательно, это все черепа долихоцефальные, а вследствие малой высоты и платицефальные (ср. фиг. 4, 5, 6, 7, 10 и 11). Что касается емкости неандертальского черепа, то первоначально она определялась (вследствие неполной целости черепа) различно, но вероятнее, что она не превышала 1230 кв. см, как согласно определяют Гексли и Швальбе, и след., приближалась к объему черепов низших рас. Близки к черепам H. neanderthalensis остатки, найденные Крамбергером в Кроации около Крапины. Выступание надбровных дуг на этих черепах еще сильнее выражено, чем на неандертальском черепе, и сильнее, чем у яванской ископаемой обезьяны Pithecantropus (см. Узконосые обезьяны), но зато лоб, по-видимому, не представлял такого отклонения кзади, а стоял вертикально, хотя по найденным обломкам судить об этих остатках с точностью нельзя. Вообще же говоря, между делювиальными черепами можно различать уже несколько рас. Так, французские антропологи различают: канштадтскую, фюрфоозскую, или гренельскую, и кроманьонскую расы. К канштадтской расе, получившей свое название по обломку черепа, найденного в 1700 г. близ Канштадта в Вюртемберге и характеризующейся сильным выступанием надбровных дуг и долихоцефалией, относили и неандертальский череп. Раса фюрфоозская, или гренельская (остатки найдены близ Фюрфооза в провинции Намюр около Гренелля), брахицефалична, тогда как раса кроманьонская — долихоцефалична. К ней относится найденный Шмерлингом 1855 г. в пещере Энгис (около Льежа) череп с тремя скелетами, получивший название энгизского, а равно черепа и остатки кроманьонской пещеры, около Ложери, найденные в 1868 г. Эти последние черепа по емкости (от 1490 до 1590 кв. см, может быть и более) превышали средний череп современных парижан (1558). Кроме перечисленных, известен целый ряд делювиальных остатков, носящих названия по местам их нахождения (Солютре, Мадлен, Клиши, Ольмо и др.). Важно, таким образом, отметить что долихоцефалы, брахицефалы, мезоцефалы и другие встречаемые между современными расами типы черепов были уже налицо в делювиальный период. Некоторые черты, свойственные неандертальскому черепу, свойственны черепу, найденному около Гибралтара (Farbes-Quarry), но его древность не определена, а также черепу, описанному Гами (Hamy) и найденному около Bologne-sur-Mer, а равно эгисгеймскому черепу, найденному Фоделем около Кольмара в 1865 г.; но последний череп отличается тем, что его лоб не был уклоняющимся назад и череп этот, хотя и долихоцефаличен (показатель = 74), но не был платицефаличен, а по высоте приближался к современным (высотный показатель = 56,7). Длина его 200 мм, ширина 148, высота 105. Во всяком случае этот череп уже сильно отличался от неандертальского, тогда как череп из Bologne-sur-Меr долихо-платицефаличен, с характерным выступанием надбровных дуг, а гибралтарский череп отличался и сильным прогнатизмом. Заслуживают особого внимания остатки, найденные около Сантоса в Бразилии бр. Неринг (или череп — Самбаки. Sambaquis — раковинные кучи). Низкий, уклоняющийся кзади лоб, сильный прогнатизм и особенно резко выраженное сужение черепа в височной (посторбитальной) области, столь характерное, по Вирхову, для черепов обезьян, и Pithecantropus (фиг. 8 и 9). Лоб очень узкий: у Pithecantropus ширина лба 90 — 94, но череп самбаки имеет 88 — 92 мм (смотря по способу измерения). Наибольшая длина 183, наибольшая ширина 143, но она лежит не на теменных буграх, как обыкновенно, а позади их, а на буграх ширина = 135 мм, так что показатель черепа = или 73,8 или 77,6, смотря по тому, которую ширину брать для его вычисления. Означенное сужение черепа в височной области Неринг находит и на южн. американских черепах, так что это собственно расовый признак. Древность этого черепа осталась неточно определенной, но с ним найдены кости делювиальных животных. Таким образом, отдельные черты черепа H. neanderthalensis мы находим, в целом ряде делювиальных черепов, а равно и в современных. Весьма существенные результаты получены при изучении нижних челюстей. Из них наибольшую важность имеют челюсть ноллетская, найденная Дюпоном (1860) в Бельгии около Фюрфооз, и челюсти, найденные Ванкелем и проф. Машка (Maschka) на Шипке и около Предмоста. Ноллетская челюсть замечательна по форме нижнего угла (фиг. 13). В то время как нижний угол челюстей современных рас представляет более или менее значительный выступ (подбородок) и образует острый угол, у обезьян этого выступа нет и угол является тупым. Ноллетская челюсть имеет угол, хотя и близкий к прямому, но тупой (94°), и в этом отношении к ней приближаются челюсть черепа Спи и челюсти некоторых низших рас (жители Новогибридских о-вов). Вообще по своим пропорциям ноллетская челюсть не подходит ни к обезьяньей, ни к человеческой, как показывают след. цифры.
| Высота | Ширина | Показатель при высоте = 100 | |
| 10 парижан 10 новокаледонцев 10 негров (Африка) 4 оранга 4 гориллы | 31,2 32,9 31,8 43,7 42 | 12,7 13,8 13,4 22,2 21,5 | 40,8 40,9 42,1 50,8 50,3 |
| Ноллетская, смотря по месту измерения | 31 42 23 | 14 14,5 16 | 57,7 |
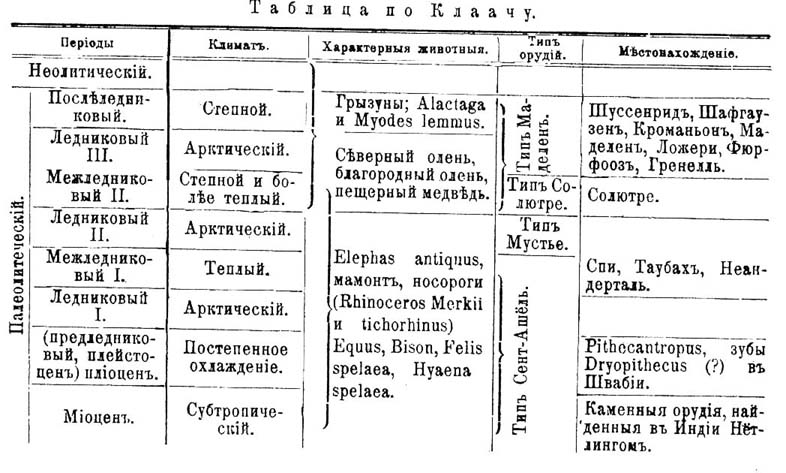
Даже Топинар склонялся признать эту челюсть за ненормальную. Челюсть шипкинская по размерам могла бы принадлежать взрослому Ч., но по зубам — является детской. Вирхов счел ее за челюсть взрослого, аномально удержавшую зубное вооружение ребенка. Во всяком случае по отсутствию подбородка, по развитию кости в толщину и ширину, по ширине основной поверхности, по присутствию ямки на месте так назыв. spina mentalis (на внутренней поверхности переднего угла) — эта челюсть является весьма своеобразной. Валькгофф, исследовавший эту челюсть при помощи Рентгеновских лучей, показал, что шипкинская челюсть несомненно принадлежит 10-летнему ребенку: внутренние полости резцов и клыков еще широко раскрыты и корни их еще не сформированы, а равно и ложнокоренные стоят на той же степени развития, как у 10-летнего ребенка. Исследованная тем же путем челюсть из Предмоста оказалась принадлежащей 7-летнему ребенку. По своим анатомическим признакам эта челюсть стоит между ноллетскою и современной, а шипкинская является более древней, чем ноллетская. Величину этих челюстей объясняют тем, что вообще челюстной аппарат первобытного Ч. достигал гораздо большего развития и относительно был больше у ребенка. Что касается до зубов, то выступание их вперед (альвеолярный прогнатизм) наблюдается на челюсти Spy № 1, хотя челюсти шипкинская и ноллетская ортогнатичны. Вообще о зубах первобытного Ч. мы знаем мало. Однако на коренных зубах, найденных в Таубахе около Веймара, Неринг описывает такие же морщинки и складочки на жевательной поверхности, какие наблюдаются у человекообразных (см.) обезьян. Такие же морщинки наблюдаются и на зубах черепов из Крапины (см. выше). Зубы из Таубаха представляют некоторые другие частности, напоминающие шимпанзе, а равно сходство с найденными Бранко в Швабии зубами, которые он приписывает ископаемой обезьяне Dryopithecus (см. Узконосые обезьяны), а Клаач считает за человеческие Что же касается прочих частей скелета, то мы остановимся лишь на конечностях. Особенно интересные данные получены Клаачем при исследовании остатков неандертальских и Spy. Бедро неуклюжей формы, а его диафиз над коленным сочленением сильно сужен и резко переходит в широкий эпифиз; суставная поверхность для надколенной чашки глубока; linea obliqua отсутствует. В отношении отсутствия lineae obliquae, глубины сочленения для чашки и др. скелет H. neanderth. напоминает скелет 3-летнего ребенка. Тонкость диафиза — наблюдается у веддов и других низших рас. Некоторые из означенных особенностей, а равно форма сочленения подвздошной кости с крестцовой — заставляют думать, что Н. neanderth. не вполне обладал способностью к вертикальному хождению. Это же выражается, по Фрэпону, на форме сочленения костей голени, а именно большой берцовой кости (фиг. 2). Судя по положению этих поверхностей, H. neanderth. ходил, имея ноги согнутыми в колене и вообще, может быть, охотнее лазал, чем ходил. Большая берцовая кость многих делювиальных рас была платикнемична, т. е. сплюснута с боков. У Н. neanderth. эта особенность слабо выражена, а именно задняя поверхность этой кости на разрезе образует выступ. Плечевая кость Н. neanderth. имела головку, направленную назад, как у низших рас, а лучевая была изогнута слегка, как у обезьян и полуобезьян (фиг. 14). В общем, это была настоящая человеческая рука. В скелете Н. neanderth. Клаач находит, однако, такие признаки, которых нет ни у Н. sapiens, ни у обезьян, и потому считает H. neanderth. не за промежуточную форму, а за форму, несколько уклонившуюся в сторону. Впрочем, мы вообще редко можем указать прямых предков для большинства животных. Что касается до распределения делювиальных остатков Ч. по периодам и их соотношения с ископаемыми остатками животных, то для выяснения этих отношений даем таблицу Мортилье и новейшую таблицу Клаача (обе в упрощенном виде).
Таблица Мортилье.
| Неолитический | Орудия типа Робенгауза (полированные каменные топоры и зазубренные кремневые стрелы) | Свайные постройки и дольмены | Климат современный | Домашние животные; сильно смешанные долихоцефалические расы | |
| Палеолитический период. | Каменные и костяные орудия | Орудия типа Маделен (зазубренные стрелы, костяные и кремневые ножи). | Пещеры, убежища и становища на открытом воздухе.
| Послеледниковый период | Северный олень; дикие быки (Bison priscus и Bos primigenius) в большом количестве; мамонт, гиены и большие кошки. Человеческие расы брахи- и мезоцефальные, приближающиеся к современным (Кроманьон и др.). |
| Орудия типа Солютре (кремневые наконечники, приостренные с обеих сторон) | Климат холодный и сухой. | |||
| Орудия типа Мустье (кремневые наконечники, приостренные с одной стороны) | Ледниковый период. Климат холодный и сырой. | Большое количество пещерного медведя, носороги. Длинноголовые низшие расы (Энгис, Ольмо и др.) | |||
| Орудия типа Сент-Ашель (большие каменные орудия миндалевидной формы). | Доледниковый период. Климат умерен. | Гиппопотам, Elephas antiquus; наиболее низкие человеческие расы (неандертальская, эгисгеймская, ноллетская и др.). | |||
Линней характеризует род Ноmо его вертикальным положением тела и менструацией. Последняя свойственна и некоторым обезьянам, а строго вертикальное положение, как мы видели, не было, может быть, свойственно некоторым делювиальным расам, и вообще для отличия видов удобнее опираться на морфологические, а не на физиологические признаки. При отсутствии защечных мешков, седалищных мозолей, гортанных мешков и os penis Ч. характеризуется короткими не достающими до колен руками, отсутствием у взрослого os centrale в кисти, варьирующим от брахи- до долихоцефалии черепом и большими пальцами ног, не противополагающимися прочим. Приводим таблицу главных пропорций тела человека и человекообразных обезьян (по Ранке); длина всего тела принимается = 100.
| Горилла | Шимпанзе | Оранг | Негры по Гульду | Южно-германские скелеты | |
| Длина туловища " руки и кисти " ноги Отношение туловища к ноге " руки к ноге Окружность головы, мм | 50,4 64,9 34,9 69,2 53,8 340 | 44,8 67,67 35,20 78,50 52,00 — | 44,50 80,72 34,72 78,00 43,30 320 | 36,98 45,16 48,47 131,00 107,30 510 | 36,27 45,43 48,83 134,60 107,40 550 |
У Ч. туловище короче, а у обезьян длиннее ноги; у Ч. рука с кистью короче, у обезьян — длиннее нога; точно так же при измерениях на скелете рука без кисти у Ч. короче, а у обезьяны длиннее ноги без стопы. Плечевая кость у Ч. короче, у обезьян длиннее бедренной. Предплечье у Ч. короче, у обезьян длиннее берцовой кости. Одно из важных отличий черепа Ч. от обезьяньего — это его емкость. Средняя вместимость черепа у европейцев, колеблющаяся около 1400 — 1500 куб. см, у низших рас такова; у бушменов ♂ (мужчин) — 1324, ♀ (женщин) — 1216; у австралийцев ♂ — 1310, ♀ — 1154; у андаманов ♂ — 1281, ♀ — 1148; у веддов ♂ — 1224, ♀ — 1151. Впрочем, minimum, наблюдавшийся у веддов, достигает 1012 куб. см, а у одной ♀ (случай Флоуэра) наблюдались 960 куб. см. Точно так же Тернер нашел 3 черепа австралиянок вместимостью менее 1000, и, по Мортону, вместимость черепа у краснокожих падает до 983. В аномальных случаях (микроцефалия) вместимость падает ниже 1000 и у европейцев. Средняя вместимость черепа человекообразных (см.) обезьян варьирует у ♂ 420 — 480 куб. см и у ♀ 380 — 410 куб. см, а у гориллы ♂ достигает 590 куб. см. Только Pithecantropus, по Дебуа, имел 1000, или, как он определил позже, 900 куб. см, и, таким образом, вместе с минимальными по емкости черепами веддов и др. он заполняет пробел между черепом Ч. и человекообразных обезьян. Из приведенных цифр видно, что вместимость черепа мужчин более вместимости черепа женщин, как это наблюдается у ♂ и ♀ обезьян. Показатель черепа определяется при длине, принимаемой = 100, и тогда, если ширина не превышает 75, мы называем череп долихоцефальным, если она колеблется от 75 до 79,9 — мезоцефальным, и, если превышает 80 — то брахицефальным. В то время, как в Австралии и Полинезии преобладают долихоцефалы, точно так же (хотя слабее) и в Африке, — в Азии и Америке (кроме долихоцефальных эскимосов) преобладание на стороне брахицефалов. Впрочем, в Азии едва ли не являются преобладающими мезоцефалы. В современной Европе длинноголовые сохранились в Скандинавии, северной Германии, Великобритании, на Пиренейском п-ове, Италии, Греции и южноевропейских о-вах и удержались на севере России, a весь центр Европы занят короткоголовыми племенами. Длинноголовое племя, найденное в наших курганах, представляет более древних обитателей Европы, которые были оттеснены наплывом короткоголовых племен из Азии при переселении народов. Форма черепа является довольно стойкой. В Скандинавии, по Ретциусу, даже теперь насчитывают не более 7% короткоголовых; такой же умеренный % был и в бронзовый, и каменный век. Человекообразные (см.) обезьяны также являются брахицефалами (оранг) и долихоцефалами (горилла и шимпанзе). Развитие надбровных дуг и сужение черепа в височной области, характерное для обезьяньих черепов, выражены, как мы видели, на делювиальных и некоторых современных черепах. Выступание нижней части лица (челюстей), или прогнатизм, гораздо сильнее у человекообразных обезьян (см. Угол лицевой), но он свойственен и низшим расам, хотя в этом отношении надо заметить, что между ними встречаются как прогнатичные, так и ортогнатичные расы. Для определения так назыв. челюстного показателя берут цифру, выражающую расстояние от переднего края затылочного отверстия до верхнего (альвеолярного) края резцов, делят ее на цифру, выражающую расстояние от переднего края затылочного отверстия до средины лобно-носового шва, и частное умножают на 100. С показателем менее 98 будут ортогнаты, с показателем 98 — 103 — мезогнаты, а с показателем выше 103 — прогнаты. В то время как у европейцев показатель 96,2, у весьма низкой расы — веддов — мы имеем еще резче выраженный ортогнатизм, 95,2. С другой стороны, более высоко стоящие соседи веддов — тамилы имеют показатель 97,7, а сингалезы — 99,2, т. е. являются менее ортогнатичными. То же самое в Африке — низко стоящие андаманы имеют показатель 101,4 (♂) и 102,2 (♀); бушмены — 96,7 (♂) и 97,8 (♀), а более высоко стоящие негры имеют показатель 104,4, т. е. более прогнатичны. Австралийцы имеют 100,6 (♂) и 99,7 (♀). Таким образом, надо допустить, что или многие низшие расы рано утеряли прогнатизм, или, наоборот, что ближайшие предки Ч. не были прогнатичны, и некоторые низшие расы вторично приобрели его. Высота хоан также представляет последовательный ряд. Европейцы имеют 24,2 мм (22 — 28), австралийцы — 24,8, негры — 23,4, готтентоты и тасманийцы — 22,8, кафры — 22,2, ведды — 19,2 (14 — 21,8), шимпанзе — 15,8. Сочленение височной кости посредством особого отростка (processus frontalls), характерное для гориллы и шимпанзе, встречается в виде исключения у многих рас и % отношения более у низших рас, чем у высших. У австралийцев встречается 15,6%, андаманов — 13, у негров — 12,4, веддов — 10,5, папуасов — 7,4, малайцев — 4,8, полинезийцев — 3,3, европейцев — 1,6. Между обезьянами: у гориллы — 100%, у шимпанзе — 88,9, у макак и циноцефалов — 80, у оранга — 29, гиббонов — 12,5. Иногда этот отросток обособляется в виде самостоятельной кости (os epipterygium); У веддов наблюдается 34,3%, у андаманов 17,4% таких черепов. В общем у веддов сочленение височной кости с лобной посредством отростка или самостоятельной кости встречается на 44,8 % черепов. Самостоятельная кость между теменными и затылочной (одиночная, парная или тройная), носящая название os incae (s. interparietale), встречается на перувианских черепах в числе 5 — 6% и гораздо реже y европейцев, у антропоморфных почти не встречается. Разделение теменной кости швом на две части встречается как у Ч., так и у обезьян (в том числе у гориллы и шимпанзе) и у последних чаще (Ранке). Присутствие подбородка на нижней челюсти, отличающее Ч. от человекообразных обезьян, сглаживается, как мы видели, отсутствием подбородка у ископаемых рас и его слабым развитием у некоторых современных. Зубы Ч. отличаются от зубов антропоморфных тем, что клыки Ч. не выдаются, на верхней челюсти нет промежутка (diastema) для помещения нижнего клыка; резцы и клыки с гладкой задней поверхностью, а равно и коренные без морщинок. Но мы видели, что, с одной стороны, у доисторических рас встречаются коренные с морщинистыми жевательными поверхностями, а с другой, зубы Dryopithecus и вообще гиббонов весьма близки к человеческим. У шимпанзе и оранга коренные зубы имеют наклонность к образованию добавочных бугорков, а обыкновенно число бугорков на верхних 4, а на нижних 5. У Ч. верхние коренные редко представляют наклонность к увеличению бугорков, но часто наклонны к уменьшению числа бугорков, а именно у европейцев и эскимосов число бугорков на 1-й, 2-й и 3-й коренном зубе выражается цифрами: 4 — 3½ — 3 или 4 — 3 — 3; у венгров, австралийцев и малайцев — 4 — 4 — 4. На нижней челюсти пятибугорчатый тип удерживают 1-й и 3-й коренной, а 2-й имеет только 4 бугорка; реже зубы трехбугорчаты и только в виде исключения 6-бугорчаты. По Цукеркандлю, Ч. свойственен зачаток зуба, сидящий позади зуба мудрости, но он прорезывается только в виде аномалии. Такая же аномалия наблюдается на черепах оранга и гориллы. Таким образом, предки приматов имели 36 зубов, подобно широконосым обезьянам. Особенности позвоночника Ч. состоят в том, что у него 12 грудных позвонков и 12 пар ребер, 5 поясничных и 5 крестцовых, тогда как у человекообразных обезьян грудных позвонков 13 и 13 пар ребер, кроме оранга, у которого, хотя и встречается то же отношение, но чаще бывает 12 грудных позвонков и 12 пар ребер, как у Ч., а поясничных у человекообразных 4, крестцовых тоже 5. У гиббонов при 13 грудных, 5 поясничных, но зато 4 крестцовых. Сравнивая эти числа, можно видеть ясно выраженную тенденцию к перемещению крестца вверх, достигающую наибольшего выражения у Ч. в тех случаях, когда (в виде аномалии) в состав крестца входит и последний поясничный позвонок. В общем, позвоночник человекообразных отличается от такового высших рас Ч. отсутствием поясничного изгиба. У высших рас позвоночник в поясничной области спереди выгнут, а у человекообразных обезьян и также у низших рас вогнут. Если взять сумму длиннот поясничных позвонков на переднем и на заднем крае, то она будет неодинакова: при выгнутой вперед поясничной области сумма переднего края будет больше, а при вогнутой — сумма заднего будет больше. Если сумму длиннот переднего края считать равной 100, то отношение суммы длиннот заднего края к ней определится следующим образом:
| У мужчин | У женщин | |
| Шимпанзе Австралийцы Андаманы Негры Ведды Европейцы | 121,2 110,1 106,2 106,0 103,3 96,2 | 112,7 103,1 102,4 103,4 99,9 93,5 |
Половое отличие этой особенности, на которую обратили внимание Тернер и Куннингам, в том, что у женщин и самок антропоморфных выгнутость и вогнутость переднего края слабее. Остистые отростки на средних шейных позвонках Ч. развиты слабее и раздвоены; у человекообразных они развиты сильнее, особенно громадны у гориллы (в зависимости от развития связок, поддерживающих голову) и не раздвоены. Раздвоение слабо выражено у готтентотов и др. Характерные отличия
| "БРОКГАУЗ И ЕФРОН" >> "Ч" >> "ЧЕ" >> "ЧЕЛ" |
Статья про "Человек, зоологический очерк" в словаре Брокгауза и Ефрона была прочитана 1146 раз
| Бургер двойного помола |
| Ананасы на гриле |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел