
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Рыбы
Определение "Рыбы " в словаре Брокгауза и Ефрона
Рыбы
Рыбы *
Содержание: Состав класса. — Форма тела. — Накожные покровы. — Органы свечения . — Мускулатура. — Скелет. — Нервная система. — Органы чувств. — Органы пищеварения. — Органы дыхания и придаточные органы кишечника. — Кровеносная система. — Мочеполовая система. — Развитие. — Географическое распространение. — Ископаемые формы. — Классификация.
Р. (Pisces) — позвоночные животные с конечностями в виде плавней, двухкамерным сердцем, жаберным дыханием и непостоянной температурой крови.
Состав класса: класс Р. заключает в себе отряды: 1) Cyclostomi, или круглоротые, противопоставляемые иногда не только всем прочим рыбам, но и всем позвоночным по отсутствию челюстных костей (бесчелюстные, Agnatha, в противоположность прочим челюстноротым позвоночным, или Gnathostomi) и по присутствию одного непарного обонятельного органа (Monorhyni, в противоположность прочим позвоночным, имеющим два обонятельных органа и называемым Amphirhyni); 2) Selachii (акулы, скаты и химеры); 3) Ganoidei, или ганоидные Р.; 4) Dipnoi, или двудышащие; 5) Teleostei, или костистые. Иногда ганоидные, костистые и двудышащие соединяются вместе под именем Teleostomi, т. е. имеющих рот на конце рыла, в противоположность Selachii, имеющим рот на брюшной стороне головы.
Форма тела весьма характерна для Р. и общеизвестна. Интересно отметить, что иногда Р. имеют почти шарообразную форму, иногда сплюснутую в спинно-брюшном направлении, как у скатов (табл. IX, фиг. 4), иногда сплюснутую с боков, но при этом асимметричную, так как глаза помещаются на одной (правой или левой) стороне, которая одна только пигментирована (камбалы и др. родственные им формы; см. табл. III, фиг. 12, 13 и 14) и т. п. На теле Р. всегда имеются непарные плавни и обыкновенно парные. Непарные плавни: один или два спинных, хвостовой и заднепроходный (табл. I, фиг. 1). В некоторых случаях (например, у угрей, табл. II, фиг. 3) все эти плавни сливаются в один непрерывный плавень. Плавни эти, как и парные, поддерживаются хрящевыми или костными лучами, а у селахий еще дистальными лучами, похожими на роговые. Особенного внимания заслуживает хвостовой плавень. Если спинная струна, проходя посредине, разделяет его на верхнюю и нижнюю части, равные по величине, то такой случай называется дифицеркией (Cyclostomi, Dipnoi). Если хорда, отклоняясь вверх, делит его на верхнюю и нижнюю части, из которых первая значительно больше второй, то такое явление называется гетероцеркией (скаты, некоторые ганоидные). При таком же направлении хорды, причем конец ее облекается хрящевым или костным футляром (уростилем), хвост снаружи может все-таки не представлять такой разницы в величине верхней и нижней части, вследствие того что лучи нижней половины гораздо длиннее лучей верхней. Этим различие в величине этих частей маскируется и хвост кажется как будто дифицеркным, не будучи им на самом деле. Такое явление называется гомоцеркией (костистые рыбы). Боковые плавни, представляющие гомолог передних и задних конечностей прочих позвоночных, могут отсутствовать вовсе, как, например, у круглоротых и некоторых других Р., или могут отсутствовать только задние плавни, как это имеет место в нескольких семействах костистых рыб, соединяемых иногда под именем Apodes, т. е. безногих. Задние плавни или удалены от передних (Pisces abdominales), или стоят очень близко к передним (Р. thoracici), или даже помещаются впереди передних плавней, под горлом (Р. jugulares).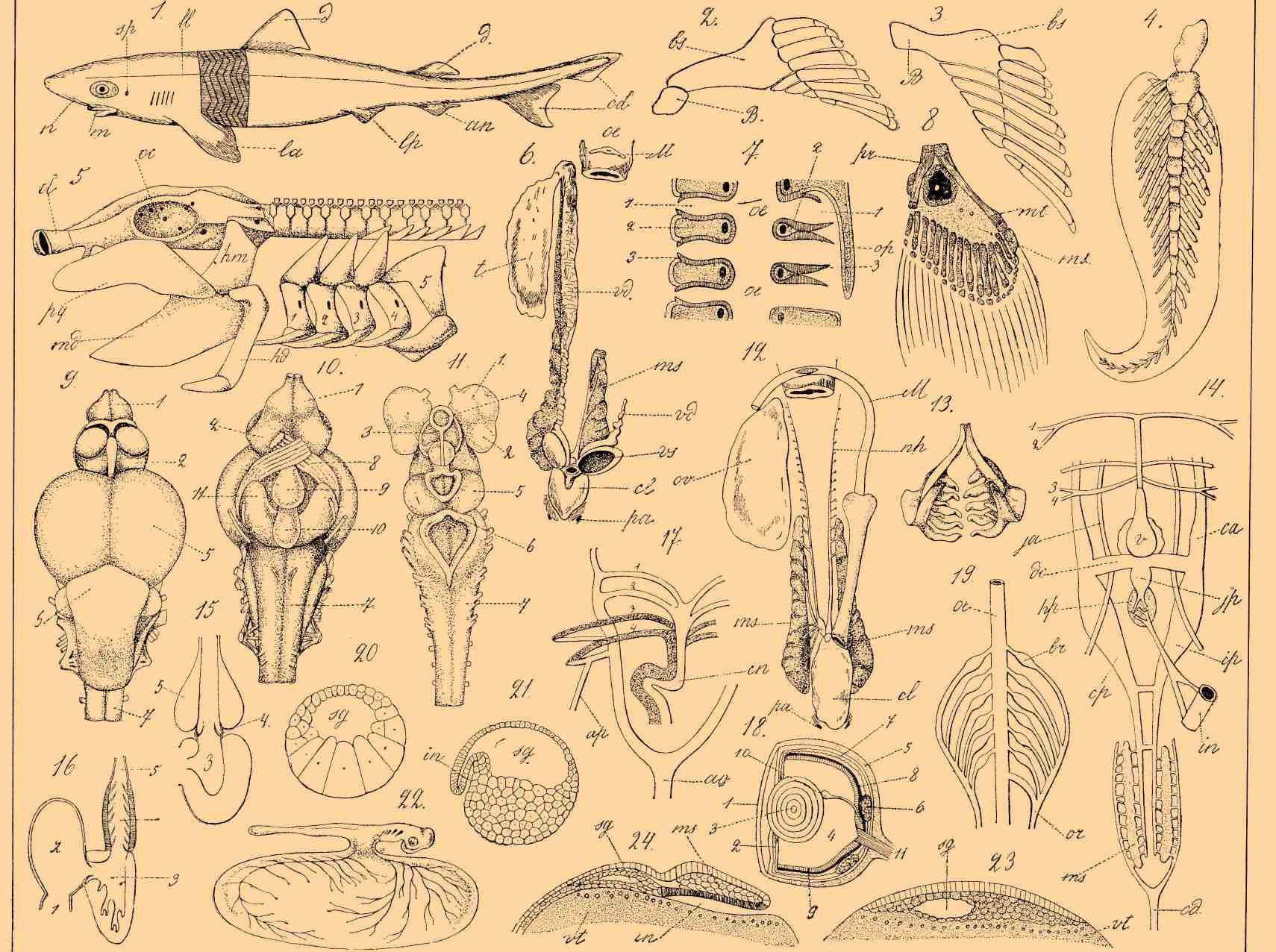
РЫБЫ I.
1. Акула Mustelus antarcticus; n — ноздри; m — рот; sp — брызгальце, la — передние боковые плавни, lp — задние боковые; d — спинные; an — заднепроходный плавень, cd — хвостовой; ll — боковая линия. Часть кожи удалена для показания расположения мышц по сегментам. 2. Задний плавень скафиринха (из ганоидных); os — слившиеся основные лучи, B — зачаток таза. 3. Задний плавень стерляди; bs — слившиеся основные лучи, B — отросток, представляющий зачаток таза и являющийся обособленным у скафиринха. 4. Плавник одной из двудышащих Р. (Ceratotus). 5. Скелет головы и передняя часть позвоночника акулы (Chiloscyllium); ol — обонятельная капсула; oc — глазница; pq — нёбно-квадратный хрящ; md — нижнечелюстной или мандибуларный; hm — гиомандибуларный; hd — гиоидный; S — жаберные дуги. 6. Мочеполовые органы самца ската; t — семенник (другой семенник удален); vd — семяпровод; ms — почки; vs — семенные пузыри; cl — клоака; pa — абдоминальные поры; M — рудимент Мюллерова канала, или яйцевода самки. 7. Схема жаберного аппарата в горизонтальном разрезе. Справа с оперкуларной крышкой — op, слева — по типу селахий; oe — пищевод: 1 — жаберная щель, 2 — жаберная дужка, или простенок между щелями; 3 — жаберные лепестки. 8. Передний плавень Polypterus; pr — propterygium, ms — mesopterygium, с окостенением посередине, mt — metapterygium. 9. Мозг лосося сверху (объяснение см. фиг. 11). 10. Он же снизу (объяснение см. фиг. 11). 11. Мозг пескоройки или личинки миноги сверху; 1 — обонятельная лопасть с отходящими обонятельными нервами, 2 — полушария переднего мозга; на рис. 9 полушария вскрыты и видны ганглиозные утолщения их нижней стенки; 3 — промежуточный мозг; 4 — теменной глаз и позади его лежащий эпифиз; 5 — средний мозг; 6 — мозжечок; 7 — продолговатый мозг с ромбоидальной ямкой у пескоройки; 8 — зрительные нервы; 9 — воронка; 10 — сосудистый мешочек (sacculus vasculosus); 11 — нижние лопасти (lobi inferiores). 12. Мочеполовой аппарат самки ската, ov — яичник; M — яйцеводы (Мюллеровы каналы) с одной общей воронкой (правый яйцевод отрезан); ms — почки; nh — почечные воронки; cl — клоака; pa — абдоминальные поры. 1 3. Нижнеглоточные кости и зубы густеры (Blicca björkna). 14. Схема венозной системы акулы, 1, 2, 3, 4 — жаберные артерии, ca — передние кардинальные вены; cp — задние кардинальные; ja — передние яремные; jр — задние яремные, слившиеся в один яремный синус; vl — боковые вены, dc — кювьеровские протоки, cd — хвостовая вена, ms — почки, hp — печень, v — сердце, in — отрезок кишки. 15. Сердце костистой Р. (объяснение см. фиг. 16). 16. Сердце акулы, схематично в разрезе; 1 — венозный синус, 2 — предсердие, 3 — желудочек, 4 — артериальный конус, 5 — truncus arteriosus. 17. Схема артериального кровообращения двудышащих; 1—4 — жаберные артерии, ap — легочная артерия, cn — артериальный конус, разделенный на две части продольной перегородкой. 18. Глаз форели в разрезе; 1 — роговица, 2 — радужина, 3 — хрусталик, 4 — стекловидное тело, 5 — склера, 6 — сосудистая оболочка с ее так называемыми железами, 7 — серебристая оболочка, 8 — пигментный слой, 9 — сетчатка, 10 — серповидный отросток, 11 — зрительные нервы. 19. Жаберный аппарат Myxine; oe — пищевод, br — жаберные мешочки, содержащие лепестки, or — открывающийся наружу общий для всех щелей канал.
20 и 21. Яйцо миноги в стадии бластулы и гаструлы, в разрезе; sg — сегментационная полость, in — вдавление для образования будущего кишечника, vt — клетки, содержащие желток. 22. Зародыш акулы с желточным пузырем. 23 и 24. Часть яйца акулы в разрезе в стадии бластулы и гаструлы; sg — сегментационная полость, in — вдавление для образования кишечной полости, ms — мезодерма, vt — желток питательный с находящимися в его верхнем слое клеточными элементами (мероцитами).
Накожные покровы Р. отличаются отсутствием рогового слоя и многоклеточных желез. Все железы, исключая железы двудышащих Р., одноклеточны и выделяют слизь. Железы двудышащих выделяют слизь, которая вместе с илом образует шар, в котором Р. переживает сухое время года. Ядоотделительные железы у некоторых Р. находятся при основании колючих лучей на парных или непарных плавнях и имеют форму многоклеточных замкнутых мешочков. Пузырьки эти лопаются лишь при надавливании, и тогда ядовитая жидкость вытекает, иногда по желобку, которым в некоторых случаях снабжен острый луч (см. Рыбный яд). В коже большинства Р. заключены чешуи. У селахий они имеют полное сходство с зубом и одеты слоем эмали (ср. Зубы и Позвоночные). Такие чешуи называются плакоидными. У прочих Р. чешуи представляют простые костные пластинки. У некоторых ганоидных Р. они имеют ромбическую форму, покрыты слоем, похожим на эмаль (ганоином), и называются ганоидными. У других Р. чешуи бывают или с округлым краем и называются циклоидными, или с гребенчатым и называются ктеноидными. Наконец, в коже могут отлагаться окостенения, как, например, у осетровых. Иногда эти окостенения одевают Р. кругом, оставляя лишь места для причленения конечностей, как это имеет место у некоторых современных Р. и ископаемых.
Органы свечения залегают в коже некоторых Р., главным образом глубоководных, и имеют, по большей части, форму глазков. Они состоят в общих чертах из утолщенной чешуи, играющей роль линзы, и лежащего под ней органа. Этот последний состоит из наружного пигментного слоя, более внутреннего, содержащего кристаллы гуанина и являющегося рефлектором, и наконец внутренней массы, производящей свет. Строение последней не вполне изучено, но иногда напоминает собой совокупность трубчатых железок.
Мускулатура у Р. сохраняет свое метамерное расположение, что ясно видно по строению 4-х продольных мышц (двух брюшных и двух спинных), составляющих главную массу тела Р. (табл. I, фиг. 1). Каждая мышца разделена на большое число сегментов поперечными прослойками соединительной ткани, называемыми миокоммами, тогда как самые мышечные сегменты, заключенные между миокоммами, называются миомерами. Одним из местных видоизменений мышечной ткани являются электрические органы (см.).
Скелет Р. представляет различные степени развития: у круглоротых он является иногда в большей своей части соединительно-тканным и лишь местами содержит хрящи, у селахии и многих ганоидных — хрящевым, а у костистых Р. является сплошь костным. Череп круглоротых, селахий и некоторых ганоидных является хрящевым, имеющим форму удлиненной коробки, заключающей в себе головной мозг и сросшейся с тремя парами хрящевых капсул для помещения органов чувств: обоняния, зрения и слуха (табл. I, фиг. 5). Снизу к черепу примыкает несколько хрящевых подвесков (табл. I, фиг. 5), имеющих форму дуг, составленных по большей части из нескольких хрящиков. Дуги эти называются висцеральными (см. Позвоночные). У ганоидных к этому хрящевому черепу прилегают кости, называемые накладными, образовавшиеся в соединительной ткани кожи или вообще прилежащих органов. Кости по большей части прилегают к черепу и легко могут быть от него отделены. В более тесную связь приходят кости, развивающиеся за счет соединительной ткани слизистой оболочки рта, например, весьма длинная парасфеноидная кость, развивающаяся на нижней стороне черепа и являющаяся единственной костью в черепной коробке у осетровых. Но у высших ганоидных, двудышащих Р., а главное у костистых — число черепных костей чрезвычайно возрастает, причем к костям соединительно-тканного происхождения присоединяются кости, развившиеся на месте хряща (эндохондрально), и все вместе они вытесняют хрящ и слагаются в весьма сложный костный череп. Верхняя часть первой висцеральной дуги представлена у низших Р. с каждой стороны нёбно-квадратным (palato-quadratum) хрящом, играющим роль верхней челюсти (табл. I, фиг. 5), а у высших особенными нёбными (palatina) и квадратными (quadrata) костями. В черепе круглоротых Р., как самостоятельный хрящ, эта дуга отсутствует, но, может быть, она представлена небольшой дужкой, лежащей под глазом и приросшей к черепу. Отсутствует у них и нижняя часть первой висцеральной дуги, представленная у прочих Р. так называемым Меккелевым хрящом или хрящевой нижней челюстью (mandibula), на месте коей у высших Р. развивается целая совокупность костей. Вторая дуга представлена в верхней части гиомандибулярным хрящом (hyomandibulare), служащим подвеском для верхней и низшей челюстей и для гиоидного хряща (hyoideum), представляющего собой нижний отдел той же второй дуги. Эти части у некоторых ганоидных и костистых Р. тоже заменяются отчасти костями, причем гиомандибулярная кость соединяется с челюстными не непосредственно, а посредством промежуточной косточки (symplecticum). Половины висцеральных дуг одной стороны соединены с половинами другой стороны при помощи непарных хрящиков (copulae). Непарный хрящ гиоидной дуги иногда достигает весьма сильного развития и служит опорой языку (os entoglossum). На гиоидной дуге развивается складка кожи, прикрывающая у высших Р. жаберные отверстия, или жаберная крышечка (operculum), a в ней развивается также комплекс костей, называемых оперкулярными. Остальные дужки, коих число у некоторых акул достигает 6 и 7, а обыкновенно бывает 5, залегают между жаберными отверстиями и называются жаберными. Непарная нижняя часть задней из них является иногда расширенной и прилегает к сердцу с нижней стороны. Верхние части жаберных дуг, будучи снабжены иногда зубами, получают название верхнеглоточных костей. Точно так же задняя (5-я) пара жаберных дуг иногда несет зубы и носит название нижнеглоточных костей (табл. I, фиг. 13). У круглоротых рыб от второй дуги с ясностью сохранилась лишь часть, соответствующая os entoglossum, a все жаберные дуги (числом 9) образовали сложную хрящевую решетку, подпирающую жаберный аппарат, а сзади заканчивающуюся капсулой, облекающей сердце. Осевой скелет туловища представлен или отдельными хрящиками, залегающими в соединительной ткани, облекающей спинную струну, сохраняющуюся у низших P. на всю жизнь и известную в продаже под именем вязиги, или же развиваются настоящие, по большей части двояковогнутые позвонки со всеми их главными частями (см. Позвонок). В позвоночнике можно отличать две части: туловищную и хвостовую. У Polypterus каждый туловищный позвонок несет две пары ребер: верхнюю и нижнюю, но у прочих Р. сохраняется лишь нижняя. Хвостовые позвонки обыкновенно снабжены нижними дугами, причем у ганоидных в состав нижних дуг входят и ребра, а у костистых ребра могут быть, одновременно с нижними дугами, на одном и том же позвонке и причленяться к ним. Иногда ребра совершенно отсутствуют. Передний пояс представлен у селахий хрящевой дужкой, к средней части коей и причленяется конечность. У ганоидных к хрящевому поясу прилегает ряд костей, не отличающихся от прочих кожных костей этих Р. У костистых Р. хрящевой пояс постепенно вытесняется костями частью накладными, частью эндохондральными. Задний пояс является в виде одного непарного хряща (например, у селахий и двудышащих), или в виде двух зачаточных хрящиков (у некоторых ганоидных), или отсутствует вовсе (у костистых и ганоидных). Исходным типом для скелета боковых плавней мы должны признать складку кожи, в которой залегают отдельные хрящевые или костные лучи. Но в такой форме у ныне живущих Р. конечность не сохранилась и простейшую форму представляет задняя конечность некоторых ганоидных, у коих скелет ее состоит из отдельных лучей первого и второго порядка (табл. I, фиг. 3), но при этом часть передних основных лучей слилась вместе и образовала более крупный хрящ (basopterigium). У других ганоидных (табл. I, фиг. 2) это слияние пошло еще дальше, причем от основного хряща отделяется особый хрящик, составляющий вместе с хрящом противоположной стороны первый зачаток заднего пояса. Если оба эти зачатка сольются, то получится непарный пояс двудышащих и др. В некоторых случаях (Polyodon) все основные лучи сливаются в один хрящ, однако, со следами расчленения. В других случаях (табл. I, фиг. 8) основные лучи, вместо того чтобы образовать один основной хрящ, могут образовать три основных части (pro-, meso- и metapterygium), кои и причленяются к поясу. У костистых Р. число этих основных частей может низводиться опять до одного (вероятно, остается metapterygium). Совершенно особое место занимают конечности двудышащих Р. (табл. I, фиг. 4), представляющая основной членистый луч, на коем с обеих сторон, или с одной, сидят боковые лучи, или их вовсе нет. На основании изучения эмбриологии и ископаемых форм должно признать, что эта конечность образовалась таким образом, что первоначально в боковой складке лежавшие отдельные лучи, вследствие возникновения и удлинения самой конечности, расположились по продольной оси плавня в два почти параллельных ряда, соответствующих боковым лучам. Эти ряды слились своими основными частями в два продольных по отношению к удлиненной оси конечности ствола, а два ствола слились в свою очередь в один, соответствующий основному лучу конечности двудышащих. У ископаемых двудышащих сохранилась двойственность основного луча. Плавни Р. могут быть приспособлены для ползания по дну (табл. V, фиг. 3), летания (табл. VI, фиг. 12) и других функций (см. Плавни).
Нервная система. Головной мозг Р. представляет следующие особенности: обонятельные лопасти сильно развиты, полушария остаются часто не разделенными и их верхняя стенка составлена лишь из одного ряда эпителиальных клеток (эпендима), не имеющих характера нервных, следовательно, эта часть мозга вовсе не является у Р. органом сложной сознательной деятельности, как у высших. На нижней стенке полушарий имеется парное ганглиозное вздутие (табл. I, фиг. 9). Эпифиз лишь в исключительных случаях, например, у миног, имеет глазоподобный придаток (табл. I, фиг. 11). Воронка образует три выступа: непарный задний или сосудистый мешочек (saccus vasculosus) и два боковых — нижние лопасти (lobi inferiores; табл. I, фиг. 10). Средний мозг хорошо развит, часто сильнее полушарий (табл. I, фиг. 9). Мозжечок или очень слабо развит, в виде небольшого утолщения переднего края ромбовидной ямки (табл. I, фиг. 11), или, наоборот, весьма сильно развит (табл. I, фиг. 9). Зрительные нервы или представляют простой перекрест (табл. I, фиг. 10), как у большинства костистых Р., или же волокна одного нерва пронизывают и смешиваются с волокнами другого, образуя настоящую хиазму, как у прочих Р. У круглоротых глазные нервы не перекрещиваются вовсе. Некоторые головные нервы, как, например, 11 пара, не обособлены, или являются, как 12 пара, нервами туловищными, а не головными — явление, которое объясняется тем обстоятельством, что у высших форм в состав головы входит большее число сегментов, чем у Р., и те сегменты, которые у высших форм являются головными, у Р. еще остаются туловищными. Наименьшее число сегментов входит в состав головы у круглоротых Р. Спинномозговые нервы каждой пары у низших Р. выходят из спинного мозга не на одном уровне, а немного отступив, как бы чередуясь правый с левым. У круглоротых чувствующие и двигательные нервы каждого сегмента не сливаются между собой, как у всех позвоночных, а остаются самостоятельными. Симпатическая система или отсутствует, или слабо развита.
Органы чувств. Обонятельный орган у круглоротых является непарным, а у Myxine открывается своим задним конусом в ротовую полость и служит для протока воды, когда Р. присасывается ртом. У других Р. обонятельный орган является парным и имеет форму двух мешочков, открывающихся каждый двумя или одним отверстием наружу. У двудышащих Р. оба эти отверстия перемещаются в ротовую полость, так что каждый обонятельный орган открывается в нее двумя отверстиями. У других Р. (р. Tetrodon) нет мешочков, а лишь обонятельные щупальца или совсем нет обонятельных органов. Глаз (табл. 1, фиг. 18) представляет ту особенность, что снаружи сосудистой оболочки залегает золотисто-серебристая оболочка (argentea), а квнутри от сосудистой — оболочка, содержащая кристаллы гуанина и обладающая способностью отражать свет (tapetum lucidum). Сосудистая оболочка, образующая иногда утолщения, изобилующие сосудистыми сплетениями, носящие неправильное название желез, вдается в стекловидное тело в виде особого серповидного отростка (processus falciformis), богатого сосудами и служащего для питания стекловидного тела, а конечное вздутие этого отростка (campanula Halleri) стоит в связи с сумкой хрусталика и играет роль при аккомодации глаза. У Myxine глаз рудиментарный. Только у немногих Р. имеются неподвижные складки, представляющие подобие век. У акул есть мигательная перепонка (см. Рудиментарные органы). Внутреннее ухо хорошо развито, но улитка зачаточная и без кортиева органа. Ни среднего, ни наружного уха нет. В эндолимфе имеются иногда достигающие громадной величины отолиты. Эмбриональное отверстие, образующееся при впячивании накожных покровов зародыша для образования внутреннего уха (см. Позвоночные), иногда остается на всю жизнь. Так, у селахий внутреннее ухо сообщается при помощи этого отверстия с наружной средой. Органы, подобные вкусовым, наблюдаются и на коже, и некоторые Р., по опытам Нагеля, обладают кожным вкусом. Совершенно своеобразным органом являются органы 6-го, неизвестного чувства. Они представляют, по большей части, сложную систему часто ветвистых каналов, изобилующих слизеотделительными клетками и содержащих особые нервные окончания. Эти каналы открываются отверстиями наружу, расположенными в виде так называемой боковой линии (табл. I, фиг. 1) на теле и на голове. Значение этих органов с точностью неизвестно (см. Позвоночные). Кроме того, на коже имеются и другие нервные окончания, имеющие значение осязательное, а у круглоротых — чувствующие клетки, также играющие роль осязательных органов и сходные с такими же клетками беспозвоночных.
Органы пищеварения. В своей передней части кишечник играет роль органов дыхания. Ротовая полость содержит обыкновенно язык и зубы, хотя последние могут иногда вовсе отсутствовать или находиться лишь в эмбриональном состоянии, как у осетровых Р. У селахий каждый зуб имеет целый ряд заместителей, последовательно заменяющих друг друга; а в других случаях потерянный зуб не заменяется, а функция его переходит на соседний. Зубы у Р. могут сидеть на всех костях, окружающих ротовую полость. Слюнные железы отсутствуют. За пищеводом обыкновенно следует расширенный отдел или желудок, иногда с довольно длинным слепым отростком, направленным назад, но иногда желудок не обособлен вовсе ни морфологически, ни гистологически, т. е. нет ни расширения, ни характерных для него желез. В начало тонкой кишки часто впадают слепые, иногда разветвленные придатки, называемые пилорическими (appendices pylorici). Кишка у Р., кроме костистых, представляет на внутренней поверхности спиральную складку, называемую спиральным клапаном. Деление кишок на отделы не выражено. Имеется печень, обыкновенно двулопастная и чаще с желчным пузырем. Имеется, хотя и не всегда, и поджелудочная железа. Последняя у некоторых Р. иногда заменена совокупностью мелких железок, что дало повод думать, будто эти Р. лишены вовсе поджелудочной железы. Кроме того, иногда имеется еще особая железа, открывающаяся в заднюю кишку и называемая ректальной. Задняя кишка открывается наружу самостоятельно у круглоротых, ганоидных и костистых Р. или же принимает в себя мочеполовые протоки и образует так называемую клоаку, как это имеет место у селахий и двудышащих. По бокам заднего прохода у селахий (табл. I, фиг. 12) имеются иногда особые поры (pori abdominales), сообщающие полость тела с наружной средой. Такие же поры описаны у некоторых костистых Р., но существование их у этих последних подвергается сомнению. У двудышащих эти поры находятся на дне особого углубления, открывающегося наружу около заднего прохода. Значение их неизвестно.
Органы дыхания и придаточные органы кишечника. Жаберные отверстия в числе 5, реже 6 или 7 пар (последнее у акул семейства Notidanidae) открываются или непосредственно наружу (селахии), или под складку кожи, помещенную на гиоидной дуге и называемую оперкулярной крышкой (табл. I, фиг. 7). У селахий жаберные щели имеют вид длинных мешков, несущих каждый на передней и задней стенке жаберные лепестки, в коих и происходит окисление крови. У ганоидных и костистых Р. простенки между щелями укорочены и низведены на степень небольших дужек, а лепестки переместились на наружный край каждой дужки (ср. обе половины фиг. 7, табл. I). Впрочем, у некоторых Р., преимущественно у способных оставлять на время воду, число щелей жаберных уменьшается, вследствие зарастания некоторых из них, или иногда лепестки — развиваются не на всех дужках. У селахий и некоторых ганоидных впереди гиоидной дуги имеется еще отдельно стоящая жаберная щель — брызгальце (spiraculum), которая у селахий несет рудиментарные жаберные лепестки. У круглоротых жаберные мешки открываются иначе: у миноги 7 жаберных отверстий снаружи, но внутренними концами они открываются не в пищевод, а в особый канал, находящийся над пищеводом и открывающийся в него спереди. Канал этот представляет остаток личиночного пищевода (см. Круглоротые). У Myxine (табл. I, фиг. 19) наружные отверстия жаберных мешков сливаются с каждой стороны в одно, а внутренние открываются в пищевод. Вода поступает в жабры Р. через рот, но у круглоротых, рот коих служит для присасывания, — через нос (миксина) или через наружные жаберные отверстия (минога), через которые и выходит обратно. Лишь у Protopterus между двудышащими в области плечевого пояса сидят небольшие кожные придаточки — наружные жабры; у других Р., главным образом у селахий, имеются наружные жабры в области жаберных щелей, но лишь в эмбриональном состоянии. Пищевод Р. снабжен на спинной стороне полым выступом, содержащим газы — плавательным пузырем (см. Пузырь плавательный), причем у двудышащих Р. пузырь этот, иногда являющийся двойным, всегда бывает ячеистым и открывается не со спинной, как у прочих, а с брюшной стороны в пищевод, играя роль легких. У многих костистых Р., покидающих водную среду, имеются под жаберной крышкой различные приспособления для дыхания вне воды. У Р. имеются, обыкновенно, парная грудная железа (Gl. thymus) и непарная или тоже парная (последнее у костистых) щитовидная (Gl. thyreoidea).
Органы кровообращения. Сердце Р. состоит из одного желудочка и одного предсердья (табл. I, фиг. 16). В последнее венозная кровь поступает из венозного синуса — резервуара, собирающего венозную кровь со всего тела. Желудочек отдает свою кровь особому расширению — артериальному конусу (conus arteriosus), снабженному у селахий, ганоидных и двудышащих Р. несколькими рядами клапанов и пульсирующему, а у костистых Р. (табл. I, фиг. 15) рудиментарному и снабженному лишь одним рядом клапанов. Из артериального конуса кровь поступает в непарный артериальный ствол (truncus arteriosus), снабженный у костистых Р. мускулистым расширением (bulbus arteriosus). От непарного ствола отходят боковые ветви, разбивающиеся на массу мелких капилляров в жаберных лепестках (табл. I, фиг. 14) и собирающихся в два ствола, идущие по спинной стороне и сливающихся в одну спинную аорту, дающую уже более мелкие артерии. Венозная система характеризуется симметричным расположением своих частей (табл. I, фиг. 14). Кровь из передней части тела приносится двумя передними кардинальными венами и двумя яремными, а сзади — тоже двумя задними кардинальными, двумя яремными, но последние бывают слиты в один синус, получающий кровь от печени. Все эти сосуды, кроме яремного синуса, с правой и левой стороны сливаются в один поперечно идущий и впадающий в венозный синус приток, называемый кювьеровским. В него же впадают сзади еще две боковые вены. Яремный синус тоже впадает в венозный, но независимо от кювьеровских протоков. Венозная кровь хвостовой вены проходит через почки по капиллярам, в них разветвляющимся, и принимается задними кардинальными венами, а венозная кровь внутренностей проходит через систему капилляров печени, а оттуда поступает в яремный синус. Таким образом, формируется воротная система почек и печени. Совершенно своеобразное видоизменение претерпевает кровеносная система у двудышащих (табл. I, фиг. 17). У них венозный синус, предсердие и артериальный конус разделены вдоль перегородками на две части: правую и левую. Правая часть синуса собирает венозную кровь, отдает его правому предсердию и оттуда после некоторого смешения с кровью левой стороны в желудочке, кровь поступает в правую сторону конуса; из этой части конуса почти непосредственно берут начало две задние жаберные дуги, причем от самой задней из них берет начало легочная артерия, несущая кровь к легкому. Левая часть синуса получает кровь из легких, передает его левому предсердию, а оттуда, после некоторого смешения с венозной кровью, поступающей в желудочек из правого предсердия, поступает в левую часть конуса, откуда берут начало еще две жаберные дуги. Таким образом, в заднюю пару дуг и, следовательно, в легкие поступает кровь наиболее венозная, а в переднюю пару — наиболее артериальная. Таким образом кровообращение двудышащих весьма близко к таковому амфибий. Лимфатическая система у Р. не имеет той самостоятельности, как у вышестоящих форм. По-видимому, одни и те же сосуды могут иногда нести то лимфу, то кровь, смотря по тому, в какой стадии находится процесс всасывания пищи. Имеется селезенка. Пульсирующие расширения в хвосте угря и Myxine (у последней два), по-видимому, содержат кровь, а не лимфу, и не могут считаться за лимфатические сердца.
Мочеполовые органы. Почки Р. помещаются на спинной стороне и имеют вид продолговатых компактных тел. Морфологически они соответствуют мезонефросу (см. Позвоночные), но передняя часть почки представляет рудиментарный остаток пронефроса. В некоторых случаях, например у селахий, не весь мезонефрос превращается в компактную массу, а часть его (табл. I, фиг. 12) имеет форму отдельно стоящих воронок. Мочеточники образуют часто расширения, служащие для собирания мочи, и открываются или в клоаку, если она имеется, или вместе с половыми протоками одним отверстием, или даже совершенно отдельно особым отверстием, лежащим позади полового. У двудышащих в клоаку открывается слепой придаток, мочевой пузырь, не имеющий непосредственной связи с мочеточниками и соответствующий таковому амфибий и других вышестоящих форм. Половые органы в редких случаях являются гермафродитными (Myxine, Serranus или морской окунь, табл. III, фиг. 4), а обыкновенно полы обособлены, причем женские половые органы представляют два типа. У одних — яичник является в виде местного набухания перитонеального слоя с каждой стороны позвоночника. Яйцеводы при этом имеют форму протоков, открывающихся в полость тела каждый своей особой воронкой или оба одним воронкообразным отверстием (табл. I, фиг. 12.). В такой форме женские половые органы свойственны селахиям, ганоидным, двудышащим и некоторым костистым Р. Но у большинства костистых Р. яичник, появляющийся у зародыша в виде складки перитонеальной стенки, своим свободным краем прирастает к наружной стенке полости тела и таким образом образуется полый мешок, на внутренней стенке коего и назревают яйца. Яйцеводы тоже прирастают к яичнику и их полость является непосредственным продолжением полости яичника. У живородящих акул яйцеводы образуют расширение, в котором и развиваются зародыши, а у живородящих костистых Р., как у бельдюги (Zoarces viviparus), развитие происходит в яичнике. Мужские органы представляют собой семенники, стоящие у селахий в непосредственной связи с передними концами мочеточников (табл. I, фиг. 6). Точно так же у ганоидных и двудышащих мочеточники играют и роль семяпроводов, но семяпроводы костистых Р. являются вполне самостоятельными от мочеточников и, может быть, соответствуют яйцеводам самки. Впрочем, и у селахий задний отдел почки имеет тоже самостоятельный мочеточник, независимый от семяпроводов. Часто яичник или семенник одной стороны недоразвивается и тогда эти органы являются непарными. Совокупительные органы являются лишь у селахий, у некоторых живородящих костистых Р. (см. Размножения органы), а обыкновенно оплодотворение внешнее. Яйца часто одеты особыми оболочками, представляющими у селахий продукт выделения особой железы, а у других Р. видоизменение так называемых фолликулярных клеток, окружающих в яичнике яйцо. У селахий хитиновая скорлупа яйца снабжена отростками для прикрепления к подводным предметам. Иногда заботу об отложенной икре принимает на себя самец (см. Размножение).
Развитие. Дробление яйца или полное, но неравномерное (круглоротые, двудышащие) и приводит к образованию бластулы (табл. I, фиг. 20) с эксцентрично лежащей сегментационной полостью (см. Дробление), или же дробление частичное (селахии, ганоидные, костистые), и приводит к образованию небольшого диска на поверхности яйца, в толще коего также наблюдается эксцентрично лежащая полость (табл. I, фиг. 23). Там, где развитие обстоятельно изучено, наблюдалось впячивание на заднем крае зародыша, служащее для образования кишечного канала (табл. I, фиг. 21 и 24). При поверхностном дроблении, одновременно с впячиванием, клетки диска обрастают желток со всех сторон так, что процесс впячивания (эмболии) и обрастания (эпиболии) совершаются одновременно (см. Позвоночные). На поверхности яйца вырисовывается мало-помалу контур будущего зародыша, и сам зародыш обособляется от массы желтка, который после вылупления зародыша оказывается подвешенным к брюшной стороне последнего в виде пузыря, называемого желточным (табл. I, фиг. 22) и уменьшающегося с возрастом по мере потребления желтка для питания тканей зародыша. Некоторые Р. имеют в зародышевом состоянии такие придатки, которых нет у них во взрослом, например наружные жабры, костные придатки на черепных костях и т. п. Асимметричные камбалы вылупляются из яйца симметричными. Таким образом, этим формам свойственно превращение. Особенно своеобразную форму имеют личинки речных угрей, ведущих в море пелагический образ жизни, и личинка миноги или пескоройка (Ammocoetes).
РЫБЫ II.
1. Щука (Esox lucius). Длина 1,3—2 м. 2. Карп (Cyprinus carpio). Длина 0,40—0,50 м. 3. Речной угорь (Anguilla vulgaris). Длина 0,75—1,5 м. 4. Макрель (Scomber scomber). Длина 0,30—0,60 м. 5. Кефаль (Mugli cephalus). Длина 0,30—0,45 м. 6. Сциена-ворон (Corvina nigra). Длина 0,20—0,40 м. 7. Форель (Salmo fario). Длина 0,40—0,92 м. 8. Усач речной (Barbus fluviatilis). Длина 0,40—0,70 м. 9. Вьюн бородатый (Cobiris barbatula). Длина около 0,12 м. 10. Пескарь речной (Gobio fluviatilis). Длина около 0,12 м.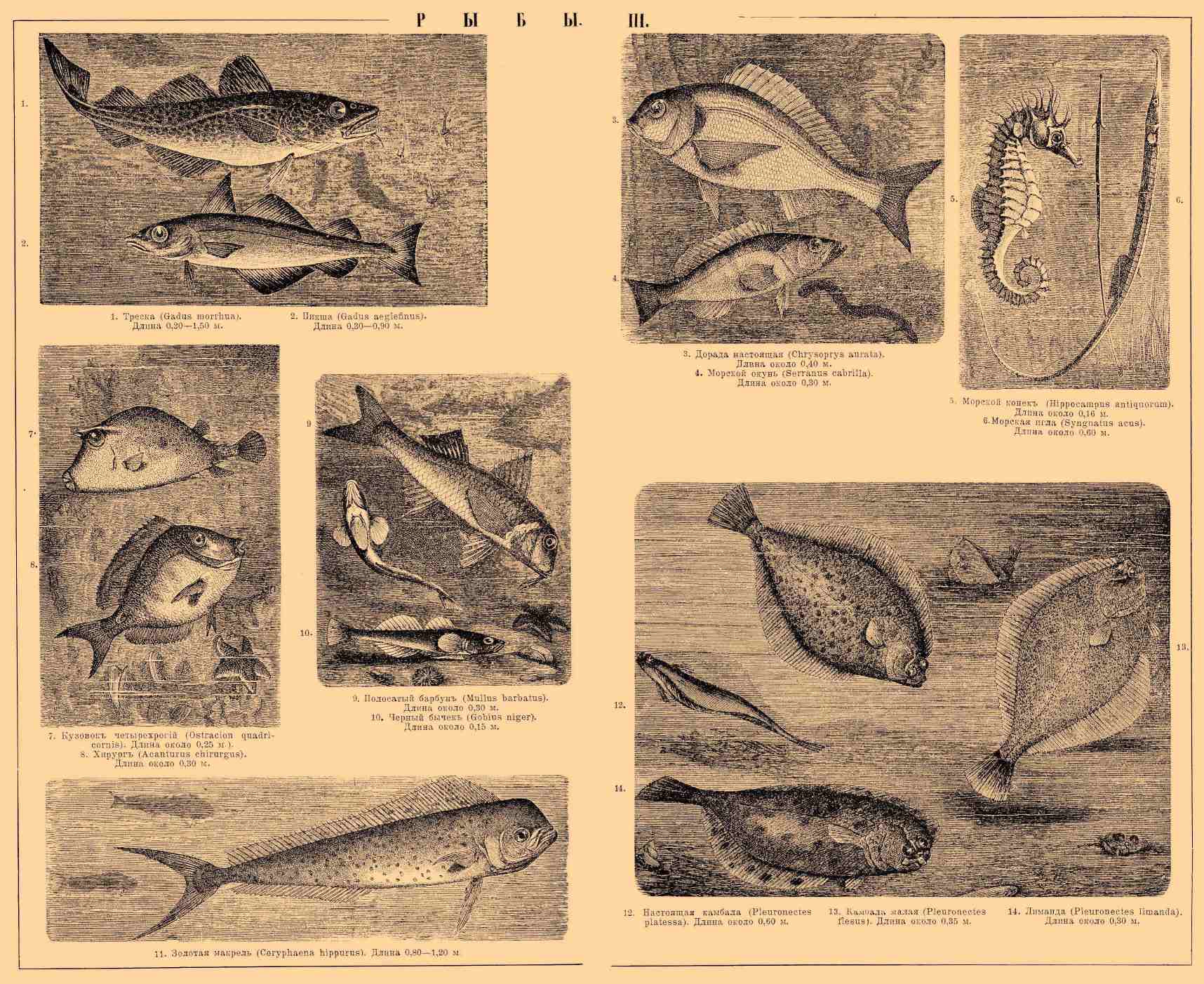
РЫБЫ III.
1. Треска (Gadus morrhua). Длина 0,20—1,5 м. 2. Пикша (Gadus aeglefinus). Длина 0,30—0,90 м. 3. Дорада настоящая (Chrysoprys aurata). Длина около 0,40 м. 4. Морской окунь (Serranus cabrilla). Длина около 0,40 м. 5. Морской конек (Hippocampus antiquorum). Длина около 0,16 м. 6. Морская игла (Syngnatus acus). Длина около 0,60 м. 7. Кузовок четырехрогий (Ostracion quadricornis). Длина около 0,25 м. 8. Хирург (Acanturus chirurgus). Длина около 0,30 м. 9. Полосатый барбун (Mullus barbatus). Длина около 0,30 м. 10. Черный бычок (Gobius niger). Длина около 0,15 м. 11. Золотая макрель (Coryphaena hippurus). Длина 0,80—1,20 м. 12. Настоящая камбала (Pleuronectes platessa). Длина около 0,60 м. 13. Камбала малая (Pleuronectes flesus). Длина около 0,35 м. 14. Лиманда (Pleuronectes limanda). Длина около 0,30 м.
Географическое распространение. Между Р. отличают: 1) исключительно морских; 2) исключительно пресноводных; 3) проходных, т. е. оставляющих море для метания икры (нереста) и идущих в реки или, наоборот, из рек в море (например, угри). Между морскими Р. отличают формы, держащиеся у берега (береговые), держащиеся в открытом море (пелагические), и глубоководные. С точки зрения распространения пресноводных Р. поверхность земли делится на три зоны: северную, характеризующуюся присутствием семейств осетровых, многочисленностью карповых (табл. II, фиг. 2, 10 и табл. VI, фиг. 1 и 2), а также присутствием лососевых (табл. II, фиг. 7) и щуковых (табл. II, фиг. 1); экваториальную зону, характеризующуюся широким развитием сомовых (табл. VII, фиг. 2); южную зону, в которой характерные для других зон семейства представлены очень бедно, но зато встречаются семейства Haplochitonidae и Galaxiidae. Северная зона обнимает неоарктическую и палеарктическую области; южная — Новую Зеландию, Тасманию и Патагонию; экваториальная — прочие части земной поверхности.
РЫБЫ IV.
1. Меченосец (Xiphias gladius). Длина 1,5—6 м. 2. Уклейка (Aspius alburnus). Длина 0,10—0,20 м. 3. Луна-рыба (Orthagoriscus mola). Длина 1,2—2 м. 4. Обыкно
| "БРОКГАУЗ И ЕФРОН" >> "Р" >> "РЫ" >> "РЫБ" |
Статья про "Рыбы " в словаре Брокгауза и Ефрона была прочитана 1274 раз
| Коптим скумбрию в коробке |
| Морской Гребешок в беконе |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел