
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Позвоночные животные
Определение "Позвоночные животные" в словаре Брокгауза и Ефрона
Позвоночные животные
Позвоночные животные*
Характеристика типа П. — Состав типа П. — Форма тела. — Накожные покровы. — Мышечная система. — Скелет. — Нервная система. — Органы чувств. — Органы пищеварения. — Кровеносная система. — Выделительная система. — Половая система. — Развитие. — Происхождение П.
П. животные (Vertebrata, Craniota, Pachycardii) хотя и получили свое название от свойственных большинству их позвонков, образующих вместе позвоночный столб, но, тем не менее, вследствие отсутствия позвонков у многих позвоночных эти органы не могут считаться характерными для этого типа. Характерным для них является: 1) форма центральной нервной системы, в виде полой трубки (фиг. 31, 42, N), лежащей на спинной стороне животного и получающей название спинного мозга, а спереди расширяющейся для образования головного мозга. Спинной мозг помещается в футляре, образованном соединительной тканью или хрящом или костными позвонками, а головной — в особом хрящевом или костном футляре — черепе. Присутствие развитого головного мозга и черепа отличает П. от бесчерепных (Acrania), почему они и получили еще название Craniota. 2) Непосредственно под нервной системой, на том месте, где у большинства П. находятся тела позвонков (см.), залегает у зародышей П. особый шнур плотной ткани — спинная струна, или chorda dorsalis (фиг. 31, 42, Ch.). Этот шнур у низших П. остается в течение всей жизни, или же удерживаются лишь участки его. Такой же шнур свойственен и оболочникам, и бесчерепным, но у бесчерепных он тянется от переднего конца тела до заднего, у зародышей оболочников он залегает лишь в задней (хвостовой) части тела, а у П. он тянется по всему туловищу и заходит лишь в заднюю часть основания черепа. Все животные его имеющие, соединяются под именем Chordata или Chordonii, т. е. хордовых. 3) На брюшной стороне П. животного помещается пульсирующее расширение кровеносной системы, состоящее из 2, 3 или 4 полостей и называемое сердцем. Этот признак также отличает П. от бесчерепных, у которых пульсирующий брюшной ствол кровеносной системы является равномерно узким (Leptocardii, или узкосердечные), почему П. иначе называются Pachycardii, или широкосердечными. 4) Передняя часть кишечного канала или в течение всей жизни, или только в зародышевом состоянии сообщается при помощи боковых отверстий или жаберных щелей с наружной средой (фиг. 35). Этот признак П. разделяют с кишечножаберными, оболочниками и бесчерепными (см. Пищеварительные органы). 5) Выделительная система у некоторых П. в течение всей жизни (у других только в зародышевом состоянии) представляет собой ряд трубок, открывающихся, подобно сегментальным органам червей, одним концом в полость тела, а другим — не непосредственно наружу, как у червей, а в два боковых канала — первично-почечных протока, а эти последние уже открываются наружу или в заднюю часть кишки, иначе в клоаку (фиг. 36). 6) Половые органы представляют местные утолщения брюшинного (перитонеального) эпителия, и для вывода половых продуктов служат различные части эмбриональной выделительной системы. В состав типа П. входят рыбы, амфибии, гады, птицы и млекопитающие. Но рыб и амфибий (представляющих большое сходство с рыбами в зародышевом состоянии) иногда соединяют под именем Ichtyopsida, а гадов и птиц, связанных целым рядом ископаемых переходных форм, соединяют под именем Sauropsida. С другой стороны, были попытки и иного подразделения П. Так, круглоротых рыб (напр. миног) на основании отсутствия мандибулярных хрящей, соответствующих нижнечелюстным хрящам других рыб, Геккель противопоставил под именем бесчелюстных, Agnatha, всем прочим П. — челюстноротым, Gnathostoma. Многими учеными рыбы и амфибии соединяются под именем Anamnia, т. е. лишенных зародышевой оболочки, называемой амнионом, а все прочие гады, птицы и млекопитающие — под именем Amniota, т. е. имеющих амнион (см. ниже). Форма тела П. животных у низших форм, живущих в воде, является весьма характерной и может быть названа рыбообразной. Но у наземных форм тело является укороченным, а хвост, если он есть, уже не является непосредственным продолжением тела, как у рыб и хвостатых амфибий, а в виде значительно более тонкого, чем само туловище, придатка. Впрочем, форма тела многих гадов, как крокодилов и ящериц, занимает середину между рыбообразной и формой тела млекопитающего. Но и между млекопитающими китообразные и Sirenia (или травоядные киты), отчасти и ластоногие (тюлени и моржи), с переходом к водному образу жизни приобретают явственно рыбообразную форму тела. У предков П., как это можно думать на основании истории развития, рыбообразное тело было снабжено плавниками, т. е. кожными оторочками, поддерживаемыми, может быть, хрящевыми лучами (фиг. 32). Плавники эти были расположены следующим образом: один плавник тянулся по всей спине, по ее срединной линии, огибал хвост и перед задним проходом раздвоялся и тянулся по бокам в виде двух складок (см. Конечность). У современных рыб непарные плавники: спинной, хвостовой и анальный представляют собой остатки этого непарного и некогда непрерывного плавня, а передние и задние боковые плавни — остатки этих боковых складок, тоже некогда непрерывных (см. также Плавники). У высших П. с переходом к наземному образу жизни исчезают непарные плавни, а начиная с амфибий, боковые плавни преобразуются в четыре пятипалые конечности, построенные уже по типу сложного рычага.
Накожные покровы у П. представляют следующие слои: 1) верхний, состоящий из многих слоев эпителиальных клеток эпидермис, который у всех П., кроме рыб, на поверхности образует роговой покров вследствие отложения в клетках поверхностных слоев эпидермиса рогового вещества, или кератина; самый верхний роговой слой обыкновенно подвергается периодическому спаданию, или линьке, или же постоянному непрерывному отшелушиванию и нарастает вновь на счет эпидермиса. Следующий слой — соединительнотканный, или corium, содержит, кроме волокон соединительной ткани, гладкие мышцы, отложения жира, пигментные клетки (хотя пигмент может отлагаться и в клетках эпидермиса), а также сосуды и нервные окончания в виде осязательных телец и т. п. Впрочем, у низших рыб, именно у круглоротых, в эпителии встречаются "чувствующие клетки" (Sinneszellen), стоящие в связи с нервами и снабженные перципирующими шипиками. Эти элементы составляют у беспозвоночных (включая и бесчерепных) общераспространенное явление и вообще представляют простейший орган чувств. На счет эпидермиса развиваются кожные железы, которые у рыб, кроме двоякодышащих, являются одноклеточными, а у этих последних, а равно и у всех прочих П., появляются многоклеточные кожные железы, особенно сильно развитые у амфибий и весьма слабо у гадов и птиц. У амфибий мы находим мешковидные, обильно выделяющие слизь железы, разбросанные по всему телу. У гадов и птиц они сосредоточены на определенных местах: так, у птиц одна сальная железа лежит над хвостом и получает название надхвостной, или копчиковой (см.). У млекопитающих находим трубчатые — потовые и гроздевидные — сальные железы, причем млечные являются у большинства млекопитающих модификацией сальных (см. Кожа). Большинство П. обладает особыми кожными образованиями. У акул и скатов в толще кожи залегают мелкие чешуйки, состоящие, подобно зубам, из верхнего слоя эмали, срединного вещества, сходного с образующим массу зуба дентином, и костной пластинки. Эти чешуйки называются плакоидными, или, иначе, кожными зубами. Сходство в строении их с зубами ротовыми говорит за то предположение, что эти последние суть не что иное, как разросшиеся и видоизменившие функцию кожные чешуйки, принадлежащие покровам, выстилающим ротовую полость. Затем чешуи прочих рыб, лишенные по большей части эмали и дентина, представляют собой, вероятно, упрощенные до степени костной пластинки плакоидные чешуйки. С другой стороны, было высказано предположение, что находимые у многих П. костные отложения в коже произошли через слияние таких же упрощенных до степени костной пластинки кожных зубов. Некоторые полагают, что перо и волос являются тоже видоизменением кожных зубов. Роговой слой принимает деятельное участие в образовании различных кожных отложений. У гадов он образует чешуи и пластинки, под коими в corium залегают часто костные отложения; у птиц он образует верхний слой пера, у млекопитающих волос, покров рогов, копыто, когти и ногти (см. эти слова) и т. п. По мнению многих, перо представляет модификацию не кожного зуба, а чешуи гадов (см. Перо), а с другой стороны, имеется ряд указаний, что зародыши млекопитающих, лишенные во взрослом состоянии кожных чешуй, имеют кожные утолщения вроде чешуй гадов, причем на каждой чешуйке появляются три волоса. Так как многие млекопитающие представляют распределение волос группами по три или такое расположение, которое может быть рассматриваемо, как видоизменение этого, то было высказано предположение, что предки млекопитающих были покрыты чешуями, причем групповое расположение волос есть указание на соотношение в их распределении с расположением чешуи (Вебер, Эмери, De-Meijere) и др. Что касается до происхождения волос, то Маурер высказал другое предположение, а именно считает его за модификацию особых органов чувств (органы 6-го чувства; см. ниже), находящихся в коже рыб и амфибий. Этому предположению противоречит, однако, то обстоятельство, что у гадов, считавшихся ближайшими предками млекопитающих, эти органы чувств отсутствуют, почему некоторые видят источник для образования волос в других органах чувств (осязательного характера), имеющих форму клеточных сосочков и сидящих на поверхности чешуи некоторых ящериц (Оппенгеймер).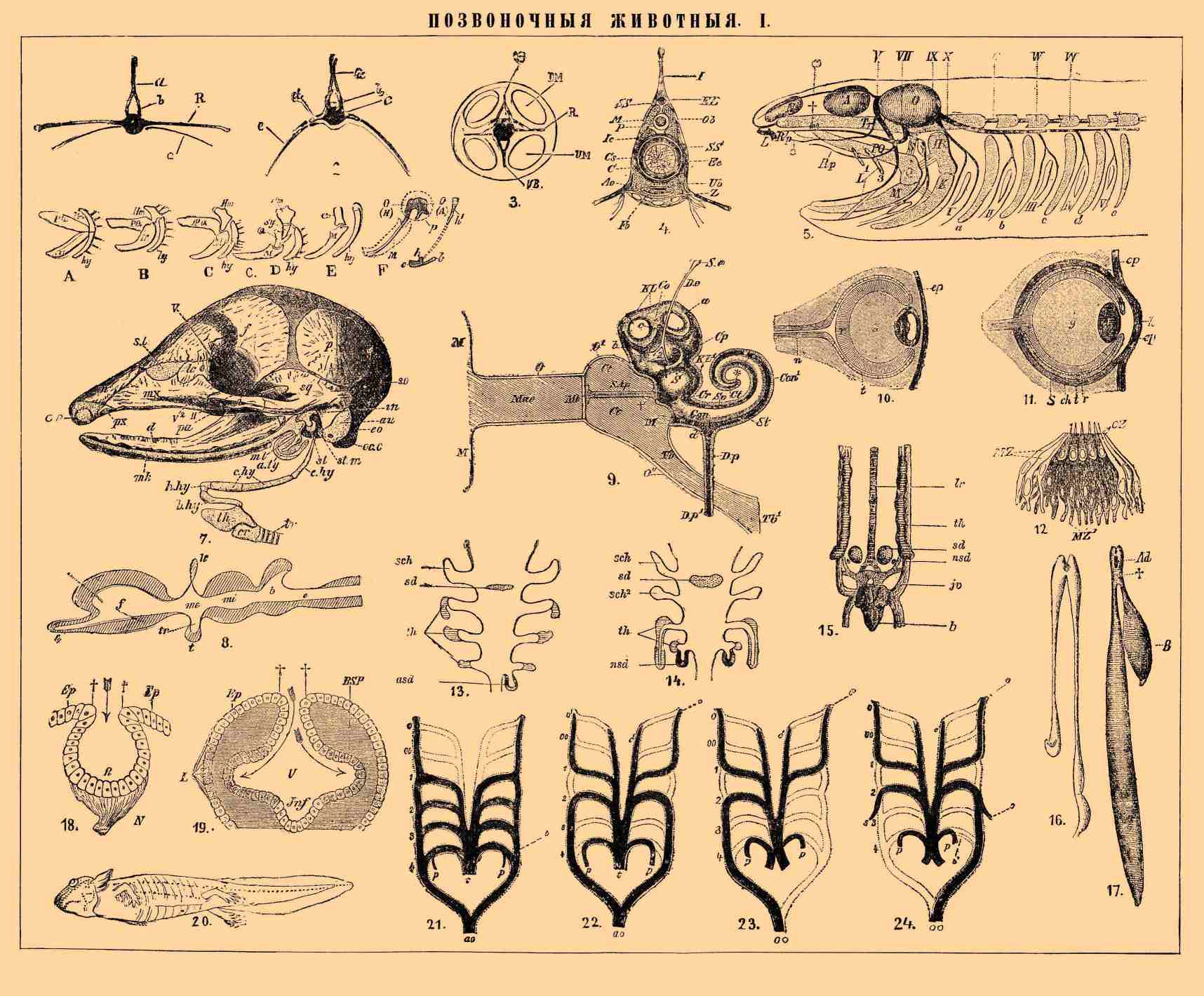
ПОЗВОНОЧНЫЕ ЖИВОТНЫЕ I.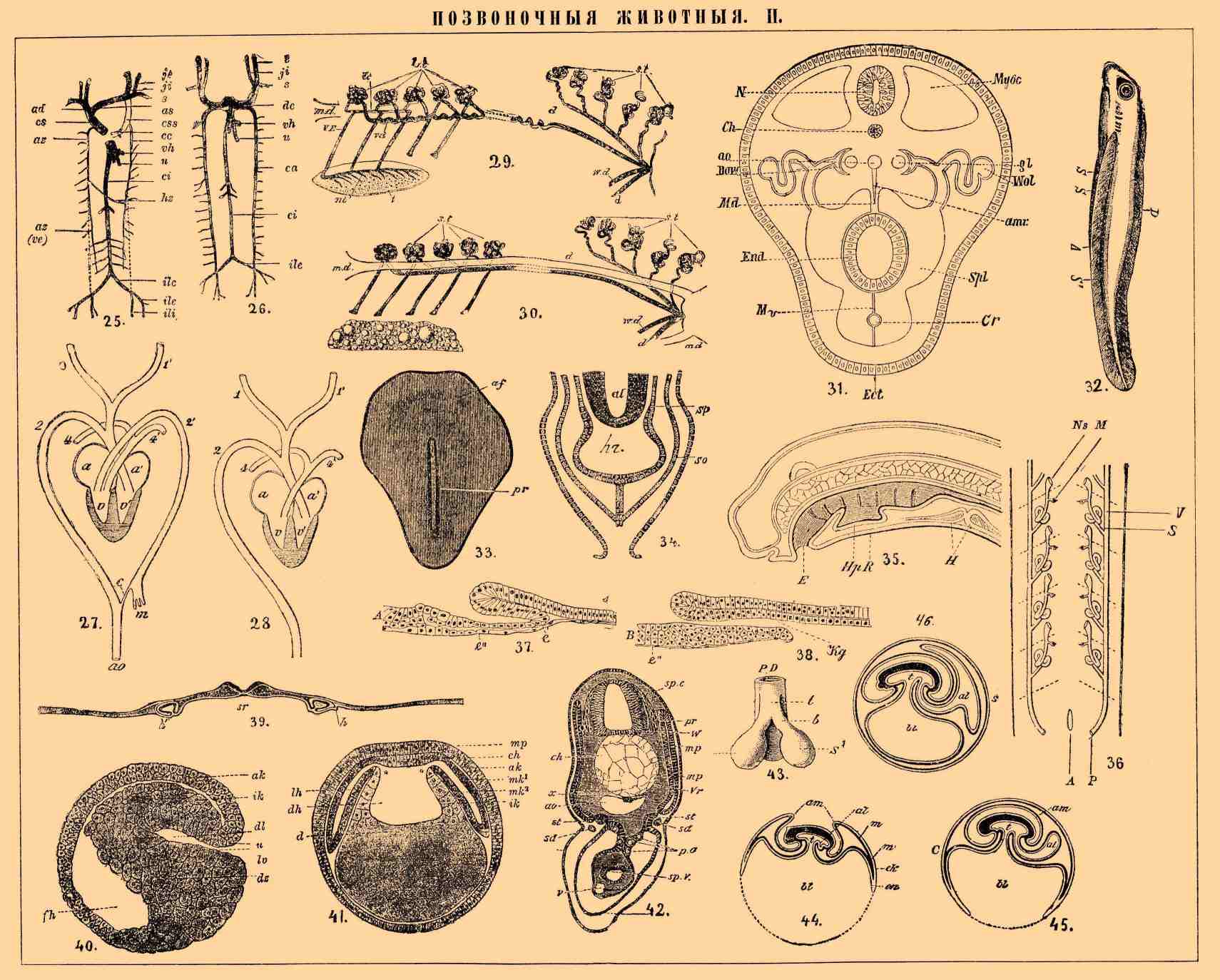
ПОЗВОНОЧНЫЕ ЖИВОТНЫЕ II.
Обьяснения к таблицам ПОЗВОНОЧНЫЕ ЖИВОТНЫЕ:
1—3. Схема позвонка и ребер: 1 — у Polypterus; 2 — у ганоидной рыбы вообще; 3 — у высших позвоночных: а — остистый отросток, b — элементы верхних дуг; c — ребра, свойственные рыбам; d — поперечные отростки; R — верхняя пара ребер у Polypterus и ребра высших П., образованные слиянием верхнего и нижнего ребра. ОВ — верхняя дуга, UВ — нижняя; DM и UM — спинные и брюшные пучки продольных мускулов.
4. Позвоночник стерляди в поперечном (по Гатчеку) разрезе; С — хорда; Cs — ее скелетогенный слой; UB, Z и FO — части хрящевой нижней дуги, облекающей аорту — ao; М — спинной мозг; P — его оболочка; ob — элементы верхней дуги; ps — остистый отросток; ER — эластическая связка над спинным мозгом.31. Схематический разрез позвоночного: Ect — экдодерма; End — энтодерма кишечника; N — нервная система; Ch — спинная струна; Муос — миоцель; Spl — спланхноцель; Md, Mv — спинной и брюшной мезентерии; Сr — сердце; Aо — нисходящая аорта с ее ветвью amr; Bow — Боуманова капсула; gl — glomerulus; Wol — Вольфов проток (из Полежаева и Шимкевича).
5. Зачаток черепа и висцерального скелета у акулы (по Видерсгейму). N, А и О — хрящевые капсулы для органов обоняния, зрения и слуха. Pr — один из парных хрящей, образующих нижнюю стенку черепа (Trabecula cranii) и считаемых некоторыми тоже за жаберную дужку; PQ и Q — части небно-квадратного хряща; М — мандибулярный хрящ; H — гиомандибулярный; К — гиоидный хрящ, L — губные хрящи, I — а, b, с, d, е — жаберные дужки; S — связка; С — хорда; W — позвонки; I—V — жаберные отверстия; VI — тройничный с его тремя главными ветвями: 1, 2 и 3; VII — личной нерв; IX — языкоглоточный; Rр — его небная ветвь; X — блуждающий; ветви блуждающего нерва и языкоглоточного так же относятся к жаберным щелям, как лицевой к лежащему между мандибулярным и гиоидной дугой брызгальцу, a тройничный — к ротовому отверстию, и именно одна ветвь иннервирует переднюю стенку, а другая — заднюю.
6. Судьба двух первых висцеральных дуг у различных П. А и В — у акул; С — у скатов; D — у костистых рыб; Ε — у гадов и птиц; F — у млекопитающих; PQ — небно-квадратный хрящ; M — мандибулярный, или Меккелев; Hm — гиомандибулярный; hу — гиоидный; Q — квадратный, у млекопитающих находящийся в полости среднего уха в виде Q и Q', или молоточка и наковальни; Sy — добавочная косточка у костистых рыб; h — часть гиоидной дуги, прирастающая в виде малого рожка к телу подъязычной — С; h' — часть гиоидной дуги, прирастающая к черепу в виде шиловидного отростка; b — большой рожок подъязычной кости, образованный следующей жаберной дужкой (см. Подъязычная кость).
7. Череп молодого броненосца (Dasypus); хрящ — затушеван (по Паркеру); — II — отверстие для выхода зрительного нерва; V и V2 — то же для ветвей тройничного; o. ty. — os tympanicum, выстилающая слуховой проход; b. hy, h. hy, с. hy, с. hу — части подъязычной кости; th — щитовидный хрящ (адамово яблоко); с ним соединены связкой малые рожки подъязычной кости; cr — перстневидный хрящ; tr — дыхательное горло; mk — мандибулярный хрящ; d — нижнечелюстная кость, на нем развивающаяся; ml — молоточек и in — наковальня, отделяющаяся от заднего конца мандибулярного хряща; st. т — стремя (гиомандибулярный хрящ); au — слуховая капсула; eo, so — затылочные кости; sq — чешуя височной, i — скуловая; p — теменная; f — лобная; tc — слезная; n — носовая; mx — верхнечелюстная; px — переднечелюстная; ра — небная; en — носовые отверстия.
8. Схема мозга П. в продольном разрезе: f — передние полушария с обонятельной лопастью — l; k — эпифиз; tr — выступ дна 3-го желудочка, или воронка (infundibulum), и под ним гипофиз — t; тe — промежуточный мозг (с 3-м желудочком); mi — средний мозг (сильвиев водопровод); b — мозжечок; e — продолговатый мозг.
9. Орган слуха человека; схема. Наружное ухо: M — ушная раковина; Мае — наружный слуховой проход; О — стенка последнего; Mt — барабанная перепонка. Среднее ухо: Ct — барабанная полость; О' — ее стенка; S Ар — слуховые косточки, представленные в виде одного палочкообразного тела; † — отвечает подножке стремени, запирающей овальное окно; M — перепонка, закрывающая круглое окно; Tb — Евстахиева труба, при Tb' открывающаяся в ротовую полость; O" — ее стенка. Внутреннее ухо (костяной футляр KL, KL' по большей части удален). S — нижний отдел слухового пузырька; а, b — полукружные каналы, из них b — срезан; Con, Con. — кожистая и костяная улитка. Se, De — остаток соединения слухового пузыря с наружной средой (ductus endolymphaticus), y акул открывающийся наружу, а у прочих П. оканчивающийся слепо; Ср — полость костного футляра, стоящая при помощи канала: d, dp, Dp' — в сообщении с лимфатическими полостями; Sv, St, Ct — полость костной улитки. Cr — канал, соединяющий верхний и нижний отдел слухового пузырька.
10—11. Две стадии развития глаза П. (из Боаса). S — склера, или белковая оболочка; h — роговая; ер — эпидерма; ch — сосудистая оболочка; i — радужина; r — сетчатка; а — ее наружный пигментный слой; n — зрительный нерв; l — хрусталик; g — стекловидное тело.
12. Комплект чувствующих клеток 6-го органа чувств в разрезе; CZ — чувствующие клетки; MZ — клетки, изолирующие их от соседних.
13—14. Две схемы развития зобной железы и щитовидной и их отношения к жаберным щелям. 13 — у зародыша ящерицы, 14 —у зародыша цыпленка (из Гертвига по де-Мерону), sch1 sch2 — первая щель; sd — щитовидная железа; nsd — ее парный придаточный отдел, td — зобная железа.
15. Щитовидная и зобная железы курицы в ее окончательном положении: sd — щитовидная железа; nsd — ее придаток, th и th1 — thymus и ее придаток; lr — дыхательное горло; h — сердце; jv — яремная вена; ca — сонная артерия (из Гертвига по де-Мерону).
16. Легкие протея (пещерной амфибии).
17. Плавательный пузырь Polypterus: Ad — вход в него из кишечника.
18. Схема развития глаза у беспозвоночных (из Видерсгейма по Каррьеру). Ep— поверхностный эпителий; R — сетчатка; N — зрительный нерв (L — клетки будущего хрусталика; BSP — клетки первичных глазных пузырей; V — полость третьего желудочка; Inf — воронка).
19. Схема развития глаза у позвоночных (из Видерсгейма по Каррьеру). Ep — поверхностный эпителий; R — сетчатка; N — зрительный нерв (L — клетки будущего хрусталика; BSP — клетки первичных глазных пузырей; V — полость третьего желудочка; Inf — воронка).
20. Боковая линия личинки саламандры (из Видерсгейма по Мальбрану).
21—24. Схема превращения артериальных дуг. 21 — двоякодышащей рыбы; 22 — хвостатой лягвы; 23 — птицы; 24 — млекопитающего (из Боаса): t — основной артериальный ствол, распадающийся на дуги 0—4; с и с 1 — сонные артерии; p — легочные артерии; b — ее соединение с корнем аорты (впоследствии зарастающее у большинства легочных), b1 — рудимент последнего у млекопитающих в виде плотного тяжа; ао — аорта; s — подключичные артерии (пунктиром обозначены части артериальной системы, существующие лишь у зародыша).
25—26. Схема развития венозной системы, 25 — более поздняя, 26 — более ранняя: ji, je — яремные (внутренние и наружные) вены или передние кардинальные; s — подключичные; vh — печеночные и пупочные; ci — нижняя полая вена; са — задние кардинальные; ili, ile — вены задних конечностей; css — остаток левой яремной; ее — венечная вена; аz — v. azydos; hy — hemizydos.
27—28. Сердце и дуги аорты крокодила (27) и птицы (28): а — правое, а' — левое предсердие; v и v1 — правый и левый желудочки; 1 и 1' — сонные артерии; 2 и 2' — правая и левая дуги аорты, причем левая делится на сосуды, отходящие к кишечнику — т, и на тонкий ствол — с 1 вместе с правой дугой, образующей нисходящую аорту — ао; 4 и 41 — легочные артерии (по Боасу). Схема.
29. Мужские мочеполовые органы зародыша акулы: t — семенник; VE — выводящие протоки его, st — сегментальные трубки; wd — Вольфов проток, при vd функционирующий как семяпровод; d — мочеточники; md — рудимент Мюллерова канала.
30. Женские мочеполовые органы зародыша акулы: ov — яичник; m. d. — Мюллеров проток; st — сегментальный трубки; w. d. — Вольфов проток. Остальные обозначения те же, что и в предыдущем рисунке (оба рисунка из Бальфура).
31. Схематический разрез позвоночного: Ect — эктодерма; End — энтодерма кишечника; N — нервная система; Ch — спинная струна; Муос — миоцель; Spl — спланхноцель; Md, Mv — спинной и брюшной мезентерии; Сr — сердце; Aо — нисходящая аорта с ее ветвью amr; Bow — Боуманова капсула; gl — glomerulus; Wol — Вольфов проток (из Полежаева и Шимкевича).
32. Гипотетический предок П. D — спинной плавень. S1 — боковые, сливающиеся позади заднего прохода А в один S1.
33. Куриный зародыш с первичной бороздой — pr и передней амниотической складкой — аf.
34. Схема образования сердца низших позвоночных: α l — кишечный канал; so — кожно-мышечный, sp — кишечно-мышечный листки мезодермы.
35. Продольный разрез через голову пескоройки. Е — вход в ротовую полость, продырявленную семью жаберными щелями; НрR — поджаберная бороздка; H — сердце (по Калберла из Гегенбаура).
36. Эмбриональная почка позвоночных; схема. А — заднепроходное отверстие; W — Вольфов проток; P — его наружное отверстие; S — сегментальные каналы; M — мальпигиево тельце; Ns — мерцательная воронка. Пунктиром обозначены границы сегментов (по Гатчеку).
37. Разрез зародыша гекко в периоде образования гастрального вдавления. Желток не изображен, е 1 — эндодерма; е" — плоские, окружающие желток клетки; Kg — отверстие бластопора в гастральную полость (из Вилля).
38. Разрез зародыша гекко после образования разрыва в дне гастральной полости. Желток не изображен, е 1 — эндодерма; е" — плоские, окружающие желток клетки; Kg — отверстие бластопора в гастральную полость (из Вилля).
39. Зародыш кролика в разрезе для показания развития сердца h и h1; sr — полость будущего кишечника; сверху — желобок — зачаток нервной системы.
40. Гаструла тритона в продольном разрезе; u — бластопор; dl — гастральная полость; fh — сегментационная полость; ak — мелкие клетки; ds — крупные, содержащие желток клетки; ik — эктодерма (из Гертвига).
41. Образование мезодермы и полости тела у тритона; схема (из Гертвига). Разрез сделан непосредственно перед бластопором: тр — нервная пластинка; ch — зачаток спинной струны; ak, ik — эктодерма и эндодерма; mk1, mk2 — кожно- и кишечно-мускульный листки мезодермы; d — желточные клетки; dh — полость кишечника, сообщающаяся при * с полостью мезодермических выступов — Ch1.
42. Разрез зародыша акулы (по Бальфуру). W, sp.c. — нервная трубка с прилежащими ганглиями — pr; ch — хорда; x — особый подхордальный шнур, тоже отделяющийся от энтодермы; ао — аорта; mp — миоцель; vr — будущий позвонок; st — сегментальная трубка в разрезе; sd — первично-почечный проток; р.о — половые клетки; sp.v — кишечник; v — вена, идущая под ним.
43. Зачаток органов дыхания П. PD — кишечник; t — трахея; b — бронхи; S1 — легкие.
44, 45, 46. Три стадии развития зародышевых оболочек птицы (по Гертвигу, схема 1); am — амнион; al — аллантоис; bl — желточный пузырь; s — сероза; m — два листка мезодермы; ek — эктодерма; en — энтодерма.
Мышечная система П., происходя у зародыша из той части среднего пласта, которая поделена на сегменты (см. ниже), у низших форм представляется явственно сегментированной. Главная масса мышц распределена в виде 4 продольных лент (2-х спинных и 2-х брюшных), причем эти ленты разделены на сегменты прослойками соединительной ткани (фасциями). Такое расположение напоминает вполне червей и бесчерепных. Но с обособлением конечностей и по мере их более сильного развития это расположение все более и более нарушается, так как расположение мышц приспособляется главным образом для движения конечностей. Таким образом, у млекопитающих можно видеть отдаленное указание на сегментацию лишь в расположении межреберных мускулов.
Скелет П. представляет собой осевую часть: череп и позвоночник с примыкающими к нему ребрами, упирающимися в грудину, и периферическую: скелет конечностей и их поясов. Главные части позвонка — его тело, верхняя дуга и нижняя (см. Позвонок) — формируются из слоя мезодермы, прилежащего к хорде и нервной системе и называемого скелетородным. У низших рыб мы видим кругом хорды и нервной системы сплошной соединительнотканный футляр, в котором залегают лишь отдельные хрящики, соответствующие тем или другим частям позвонка (фиг. 4). Хрящики, залегающие по бокам нервной трубки, соответствуют верхним дугам, а хрящики, залегающие под хордой, — нижним дугам. Во всяком случае до образования позвонков дело не доходит. У других рыб, а также и у всех вышестоящих форм в скелетородном слое образуются цилиндрические хрящи или окостенения, пронизанные хордой и представляющие собой зачаток тел позвонков. Наконец, вследствие усиления развития хряща или кости хорда может все более вытесняться. Она теряет свою непрерывность, причем остатки ее ткани находятся или между позвонками, или внутри самих позвонков, и наконец она исчезает совсем, что мы и имеем у громадного большинства П. С телами позвонков сочленяются, а у более высоко стоящих форм совершенно срастаются, дуги. Однако число дуг не всегда соответствует числу позвонков; так, напр., у некоторых палеозойских амфибий на два тела позвонка приходится лишь одна верхняя дуга, а у современных акул иногда на одно тело позвонка приходятся две и даже три дуги. К позвонкам причленяются ребра, которые отнюдь не нужно смешивать с нижними дугами, так как очень часто у рыб те и другие встречаются одновременно. У многих рыб нет вовсе ребер, а у ганоидной рыбы Роlypterus (фиг. 1) на каждый позвонок приходятся две пары ребер: верхняя и нижняя, причем последняя пара соответствует ребрам прочих рыб (фиг. 2). Что касается до ребер амфибий и всех прочих П., то их ребра, вероятно, образованы каждое слиянием двух: верхнего и нижнего, соответствующих двум ребрам Polypterus (фиг. 3). У низших П. все позвонки, кроме задних, могут нести ребра, а у высших они свойственны лишь грудным позвонкам (см.). Начиная с амфибий, появляются на груди снизу два хряща или кости: грудина (episternum) и надгрудинный хрящ или кость (episternum). У большинства П. в грудину упирается нижними концами часть ребер, а в episternum — ключицы (см. Плечевой пояс). У птиц нет episternum, а у млекопитающих, кроме ехидны и утконоса, оно частью сливается с верхушкой грудины, частью же остается в виде хрящиков, залегающих в месте сочленения ключицы с грудиной. Грудина тоже иногда отсутствует, напр. у черепах и др. Конечности (см.) рыб представляют собой плавники (см.), иногда сочленяющиеся с хрящами или костями, представляющими передний или задний пояса (см.) или же, как это часто имеет место по отношению к задним плавникам, элементы пояса могут отсутствовать, или, точнее, они еще не обособились от элементов самих плавников (см. Конечности). У пятипалых П. (Pentadaclyloidea), т. е. у всех выше рыб стоящих форм, имеются пояса конечностей, состоящие каждый из трех главных костей: передний из лопатки, ключицы и вороньей кости, причем лопатка занимает спинную сторону, а две прочие брюшную; задний пояс (образующий у высших форм вместе с крестцом таз) — из подвздошной кости, занимающей положение, соответствующее лопатке, сочленяющейся с поперечными отростками крестцовых позвонков, затем лобковой, занимающей положение ключицы, и седалищной, занимающей положение вороньей кости. Между нижними концами лобковой и седалищной кости залегают иногда ниже хрящики, но они никогда не достигают такого развития, как грудина и epistern u m. Обыкновенно же эти кости таза, сходясь своими нижними концами, образуют два сращения (symphisis). Самые конечности пятипалых форм представляют всегда следующие части: верхний ближайший к телу сегмент (плечо или бедро), содержащий одну кость, затем второй сегмент (предплечье или голень), содержащий две рядом лежащие кости; потом совокупность мелких костей, составляющих кисть и ступню, и, наконец, косточки пальцев, или фаланги. Спереди к позвоночному столбу причленяется череп, представляющий у низших форм хрящевую коробку для помещения головного мозга, и сросшиеся с ней три пары хрящевых капсул для помещения органов обоняния, зрения и слуха (фиг. 5). Но уже начиная с ганоидных рыб, к этой коробке прилегают кости, а у костистых рыб, а равно и всех вышестоящих форм большая часть хрящевых стенок коробки заменяется костями, развивающимися или из надхрящницы на месте хряща этой коробки, или из прилежащей к ней соединительной ткани (фиг. 7; см. Череп). Снизу к черепу подвешено несколько парных хрящей или костей, соединенных попарно непарными хрящиками (copulae) и носящих общее название висцеральных или жаберных или, наконец, глоточных дужек (фиг. 5), так как они залегают обыкновенно между жаберными щелями, и след., в области глотки. Впрочем, правильнее называть жаберными только задние висцеральные дужки (кроме двух передних пар), так как они действительно залегают между жаберными щелями, а у тех форм, у которых эти последние атрофированы, они соединяются с подъязычной костью (см.) или получают иное назначение. Но наибольшего внимания заслуживают две передние дуги. Первая из них образована двумя хрящами, из коих верхний носит название нёбно-квадратного (palato-quadratum), а нижний — мандибулярного, или Меккелева (фиг. 5 PQ, Q и М). У низших рыб эти хрящи, срастаясь с соответствующими хрящами противоположной стороны, играют роль верхней и нижней челюстей и ограничивают ротовое отверстие. На основании некоторых особенностей распределения нервов можно думать, что рот позвоночных есть тоже две сросшиеся жаберные щели. Начиная с костистых рыб (фиг. 6, D), небно-квадратный хрящ не играет роли верхней челюсти. Верхнечелюстные кости развиваются из соединительной ткани и замещают небно-квадратный хрящ, который представлен у этих рыб двумя костями: небной и квадратной, причем последняя служит уже подвесочной костью, к которой причленяется нижняя челюсть. Вторая дужка образована тоже двумя хрящами: верхним — гиомандибулярным, и нижним — гиоидным. Гиоидный хрящ или кость идет на образование рожков подъязычной кости (см.), а гиомандибулярный служит подвеском, к которому причленяются сначала квадратный хрящ, мандибулярный и гиоидный. Начиная с амфибий (рис. 6, Е), гиомандибулярный хрящ (или кость) помещается уже в среднем ухе и получает название columella auris, отчасти соответствующей стремени (stapes) высших позвоночных, а у млекопитающих (фиг. 7) и квадратный хрящ помещается там же, причем получает название наковальни (incus), и, кроме того, от заднего конца мандибулярного хряща отделяется еще хрящик, образующий молоточек (maleus). Таким образом формируются три слуховые косточки млекопитающих. Впрочем, вопрос о значении молоточка и наковальни некоторыми морфологами решается иначе. Подобно жаберным дужкам, гиоидные дуги правой и левой стороны соединяются между собой тоже непарным хрящиком, который вместе с непарным хрящиком следующей пары дужек и образует тело подъязычной кости, тогда как прилегающие к ней части гиоидной и последующих дужек образуют рожки (см. Подъязычная кость; фиг. 6, F). Сопоставляя все сказанное, видно, что большинство отдельных частей скелета П. животного в своем развитии проходит три стадии: первую, когда они представлены массой клеток среднего пласта или даже более специализированной соединительной тканью; вторую, когда они представлены хрящевыми элементами; третью и окончательную, когда они представлены костями. При этом надо отметить, что чем выше мы будем подниматься по животной лестнице, тем меньше будет общее число костей, составляющих скелет. Мы видели, что независимые сначала части позвонков у высших форм срастаются вместе в один позвонок; независимые и многочисленные кости черепа костистых рыб срастаются вместе и образуют небольшое число сложных костей черепа млекопитающих и в частности человека. Так, затылочная кость человека представлена у костистых рыб четырьмя отдельными костями. Еще более сложными костями в черепе человека являются кости височная, клиновидная и др. С другой стороны, жаберные дужки, все вместе представляющие так назыв. висцеральный скелет черепа, частью изменяют функцию, частью тоже входят в состав черепа, а частью атрофируются.
Нервная система состоит из центральной части, представленной головным и спинным мозгом, периферической, представленной отходящими от них нервами и находящимися в связи с ними ганглиями, и, наконец, симпатической, состоящей из ряда ганглиев, лежащих вдоль позвоночника и связанных комиссурами между собой, а также и с периферической системой. Периферические нервы, отходящие от спинного мозга, начинаются каждый двумя корнями: верхним, снабженным ганглием и чувствующим, и нижним — двигательным (см. Нервная система). Спинной мозг залегает у зародыша большинства П. в виде желобка, тянущегося по спинной срединной линии; затем, когда края желобка сойдутся, то он превращается в трубку, помещенную над накожными покровами. Передняя часть трубки образует три последовательных вздутия — три первичных мозговых пузыря. Ганглии чувствительных корешков, или спинные ганглии, возникают в виде самостоятельных утолщений эктодермы и отделяют от себя кучки клеток для образования ганглиев симпатической системы. Периферические нервы являются как выросты спинного и головного мозга или ганглиев чувствующих корешков и только впоследствии приходят в сообщение с органами, к которым они направляются. Три первичных мозговых пузыря метаморфизируются следующим образом (фиг. 8): передний дает два полых выступа — это полушария переднего мозга, содержания в себе полости, называемый боковыми желудочками (f). Полость первоначального переднего пузыря с окружающими стенками дает промежуточный мозг с третьим желудочком (те), на верхней стенке коего помещается особый придаток мозга — эпифиз (epiphysis cerebri s. glandula pinealis), а снизу полый выступ, или воронка (infundibulum), и на ее вершине другой придаток — гипофиз (hypophysis cerebri s. gl. pituitaria). Стенки второго мозгового пузыря представляют собой средний мозг (mi), а его полость низводится на степень узкого канала — сильвиева водопровода (aquaeductus Sylvii). Полость третьего пузыря остается в виде расширения, именуемого четвертым желудочком, а его стенки образуют в своей передней части мозжечок (b) и в задней — продолговатый мозг (см. Мозг). По мере поднятия вверх по животной лестнице полушария все более и более осложняются в своем развитии, увеличиваются в размерах и, загибаясь назад, соприкасаются тоже с сильно развитым мозжечком, закрывая таким образом промежуточный и средний мозг. У млекопитающих полушария становятся извилистыми. Затем у рыб и амфибий — все части мозга лежат на одной горизонтальной линии, а у прочих форм они образуют несколько изгибов, наиболее сильно выраженных у млекопитающих. От головного мозга у высших П. берут начало 12 пар нервов, но у низших П. число это менее, так как те нервы, которые у высших берут начало от головного мозга, у низших начинаются от спинного, иначе говоря, чем выше стоит П., тем большее число сегментов туловища присоединяется к голове и тем большее число спинных нервов, принадлежащих этим сегментам, становится головными. По аналогии с тем, что известно относительно туловищных нервов, морфологи давно стремятся определить, какие нервы головы соответствуют спинным или чувствующим корешкам, какие — брюшным или двигательным, и таким образом определить, скольким сегментам тела соответствует голова, так как на каждый сегмент приходится с каждой стороны по одному чувствующему и по одному двигательному корешку. Физиологический критерий оказывается неприменимым, так как многие нервы головы — смешанной природы и содержат одновременно волокна чувствующие и двигательные или же являются различной природы у различных групп. Присутствие ганглия на головном нерве также не может служить критерием, так как, с одной стороны, этот ганглий может представлять элемент симпатической системы, а с другой — отсутствие ганглия может быть позднейшим явлением. Место отхождения нерва, дающее ясное указание на его природу в спинномозговых нервах, также не может служить критерием в голове, где нервы вообще смещены на нижнюю поверхность мозга. Отсюда понятна и трудность решения вопроса. Первоначально было сделано предположение, что в голове большинство корешков как спинных, так и брюшных развилось в самостоятельные нервы, тогда как в туловище каждая спинная ветвь соединилась с брюшною для образования одного смешанного нерва. Обыкновенно за спинные корешки принимают следующие нервы: обонятельный, или olfactorius, зрительный, или n. opticus (хотя многие не считают его в числе прочих нервов, так как развитие его значительно отличается от развития других нервов; см. ниже), тройничный, или n. trigeminus (представляет результат слияния двух нервов); лицевой и слуховой представляет, по-видимому, разделившийся один нерв acustico-facialis), языкоглоточный, или glosso-pharyngoeus (вкусовой у высших П.), блуждающий (n. vagus; представляет тоже, вероятно, сложный нерв, как и тройничный). Нервы, двигающие глазами, именно oculomotoris, trochlearis и abducens, считают брюшными ветвями. Задний черепной нерв — подъязычный или hypoglossus y низших П. является настоящим туловищным нервом с спинными и брюшными
| "БРОКГАУЗ И ЕФРОН" >> "П" >> "ПО" >> "ПОЗ" |
Статья про "Позвоночные животные" в словаре Брокгауза и Ефрона была прочитана 11074 раз
| Коптим скумбрию в коробке |
| Куриный суп |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел