
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Питание растений
Определение "Питание растений" в словаре Брокгауза и Ефрона
Питание растений
Питание растений*
Характеристическая черта П. растений состоит в том, что в то время, как для П. животных нужны готовые белки, жиры и углеводы, растение само приготовляет их для себя. Пищею для растения служат простейшие минеральные соединения: углекислота, вода и калийные, известковые, магнезиальные и железные соли кислот: азотной, фосфорной и серной. Из этих простых соединений, воспринимаемых из внешней среды, растение предварительно приготовляет (синтезирует) те же белки и углеводы, и затем П. в более тесном смысле слова, т. е. образование живого вещества, совершается уже вполне аналогично тому, как это происходит и в организме животного. С этой точки зрения, в общем круговороте веществ П. растений является как бы коррективом к П. животных: последние постоянно только разрушают сложные органические соединения, выделяя их химические элементы в виде углекислоты, воды, мочевины и пр.; растение не только разрушает, подобно животному, но и снова создает их, пользуясь указанными отбросами животного обмена веществ. Эта особенность в П. резко отразилась и во всей организации растений. В то время, как животное должно было изощрять свои способности (ощущения, движения), вступать в борьбу с себе подобными для того, чтобы добыть себе пищу, для растения пища рассеяна всюду в атмосфере и в почве. Нужно было только выработать приспособления для наилучшего всасывания ее — увеличить поверхность соприкосновения с атмосферою и почвою; для этого и служит широкое развитие зеленой листвы и корней, придающее растениям их характерный вид. Далее, приготовляя само для себя необходимые питательные вещества, растение не имеет нужды и в сложных пищеварительных аппаратах. Пищеварительного акта растения, собственно говоря, не знают. Вместо сложных пищеварительных аппаратов мы находим у них сравнительно очень просто устроенные аппараты для образования (синтеза) питательных веществ (см. Растение и Ткани растительные). В этом отношении очень интересна небольшая группа насекомоядных растений (см.). Представители этой группы как будто сделали попытку питаться, подобно животному, готовою твердою пищею (насекомыми), не утратив, однако, способности синтезировать себе питательные вещества; оказывается, что именно в этой группе мы находим и чувствительность к раздражению и способность совершать быстрые движения, необходимые для ловли насекомых, и, наконец, специальные пищеварительные железки, собранные иногда в особые полости или мешки. Но насекомоядные растения составляют исключение. Типическое зеленое растение питается простейшими минеральными соединениями, само приготовляя для себя белки и углеводы. Только там, где нет хлорофилла, как, напр., в классе грибов, нужны уже готовые тройные (т. е. состоящие из углерода, водорода и кислорода) соединения; синтезируются лишь белки. П. этих растений представляет поэтому большую аналогию с П. животных.
I. Синтез питательных веществ происходит главным образом в листьях, в зеленой мякоти их. Огромное значение этих процессов в общей экономии природы станет понятным, если мы вспомним, что не только П. растений и животных всецело зависит от них, но и большая часть энергии, циркулирующей в мертвой природе, имеет своим источником эти синтетические процессы. Если исключить силу ветра и падающей воды, сравнительно мало утилизируемые, то вся остальная энергия, утилизируемая в технике, была накоплена в свое время зеленым растением.
1) Синтез тройных соединений, или усвоение углерода (С) из CO 2 и H 2 O на свету, см. Ассимиляция и Хлорофилл. К сказанному там прибавим только, что углекислотою, содержащеюся в почвенном воздухе и растворенною в почвенной воде, растения, по-видимому, не пользуются. Почвенная углекислота с током почвенной воды (см. Передвижение веществ в растении), по-видимому, не доходит до зеленой ткани растения, так как иначе растение, конечно, пользовалось бы ею точно так же, как и углекислотою атмосферы. Очевидно, что для П. растения совершенно достаточно и того количества этого вещества, которое содержится в воздухе. Окись углерода (СО), болотный газ (СН 4) и вообще все углеводороды не могут служить для П. растений. Так как продуктами усвоения углекислоты являются различные углеводы, то понятно, что если эти вещества искусственно ввести в растение, то последнее будет питаться ими. Подобных опытов П. было сделано много, причем выяснилось, что между способностью вещества служить пищею для растения и химическою структурою его существует несомненная связь. По отношению к П. растений в естественных условиях роста весьма интересным представляется вопрос, не служат ли углеродистые соединения почвы, так наз. гуминовые вещества, источником углерода для растений. Не говоря уже о грибах и бактериях, и среди высших растений есть значительная группа форм, специально приспособившихся к питанию частицами гумуса, — это так называемые сапрофитные растения (см.), имеющие особые приспособления (присоски) для растворения и высасывания разлагающихся органических веществ почвы (см. также Микорица). У большинства этих растений способность к синтезу углеводов из углекислоты сильно понижена или даже хлорофилл вовсе отсутствует. Для них, стало быть, П. гумусом не подлежит сомнению, и вопрос состоит лишь в том, не служат ли гуминовые вещества почвы источником углерода для всех вообще растений. Точного ответа на этот вопрос дать еще нельзя, что объясняется трудностью экспериментирования со столь мало изученными веществами, каковы гуминовые. Нужно думать, однако, что если гуминовые вещества и служат пищею растению, то роль их в этом отношении должна быть очень незначительна. Благотворное влияние гумуса на плодородие почвы объясняется, вероятно, косвенным участием его в акте П. растений (см. Гуминовые вещества). Растения, не имеющие хлорофилла (грибы, бактерии), не могут усваивать и углекислоты; для них нужны готовые тройные соединения. Однако в недавнее время были указаны любопытные исключения из этого общего правила. Среди бактерий есть небольшая группа форм, окрашенных в красный цвет особым пигментом — бактериопурпурином. По наблюдениям Энгельмана, эти пурпурные бактерии разлагают СО 2, подобно зеленым растениям, причем бактериопурпурин играет у них такую же роль, какую у зеленых растений — хлорофилл. Еще интереснее другой случай, указанный С. Виноградским, по исследованиям которого селитряные бактерии (т. е. бактерии, окисляющие аммиак в азотную кислоту) также разлагают углекислоту и могут поэтому развиваться в жидкостях, абсолютно лишенных органических соединений. Бактерии эти не содержат никакого пигмента и потому не могут утилизировать лучистую энергию, как зеленые растения или пурпурные бактерии. Но они окисляют аммиак и освобождающеюся при этом химическою энергией пользуются для восстановления углекислоты и образования из нее тройных соединений.
Вместе с углеродом, которого растения содержат в среднем 4%, при образовании тройных соединений усваиваются растением и химические элементы воды, т. е. водород и кислород. Первого в растениях в среднем 6,5%, второго 42%. Следовательно, растение приобретает 93,5% своего сухого вещества из воды и тех ничтожных количеств углекислоты, которые содержатся в воздухе. Остальные 6,5% сухого веса приходятся на долю азота (в среднем 1,5%) и золы (5%).
2) Усвоение азота и синтез белковых тел в растении. Для зеленых растений наиболее пригодным источником азота служит азотная кислота (HNO 3) в виде ее солей: калийной селитры, азотно-кальциевой соли и др., которые растение всасывает из почвы вместе с водою. С аммиачными солями питание идет хуже, хотя все-таки и аммиачный азот представляет хорошую пищу (Мюнц). Аммиак растения могут всасывать и через листья из воздуха. Из незеленых растений грибы и бактерии предпочитают аммиачные соли в качестве источника азота. Органический азот — мочевина, глюкокол, аспарагин и другие азотистые органические соединения, заключающие группу (NH 3), также служат превосходною пищею для растений. В естественных условиях роста растения находят указанные соединения азота в почве, куда в конце концов попадают как отбросы животного обмена веществ, так и трупы животных и растений. Здесь эти органические остатки делаются прежде всего добычею низших организмов (бактерий, грибков), которые, растворяя и разлагая их, усваивают азотистые составные части, переводя их обратно в белки своей плазмы ("регенерация белков в почве", по Костычеву). Часть азота, однако, переходит при этом в аммиак, но и этот последний недолго остается в почве; он окисляется в азотную кислоту, и это окисление совершается также под влиянием бактерий. Азотная кислота (в виде солей) поступает в почвенный раствор и частью вымывается из почвы и уносится в реки и моря, частью поглощается корнями растений. Так как, с одной стороны, в почву постоянно поступают новые количества органических остатков, а с другой — поколения микробов постоянно сменяют друг друга, причем трупы отживших поколений делаются добычею новых, то понятно, что как регенерация белков, так и образование аммиака и азотной кислоты в почве никогда не прекращаются: во всякий момент между количествами этих веществ существует подвижное равновесие. Азотистые соединения постоянно циркулируют между животными, растениями, почвою, морем и атмосферою, причем, однако, часть азота постоянно выходит из круговорота, выделяясь в свободном состоянии. Таким образом количество связанного азота должно бы постоянно уменьшаться, так как сам по себе этот элемент весьма трудно вступает в соединения; с истощением же азотистых соединений должна, конечно, прекратиться и жизнь на земле. Это соображение давно уже заставляло ученых думать, что в природе должны существовать процессы, компенсирующие указанное уменьшение количества азотистых соединений. Искомый корректив был действительно найден в последние годы, и им оказалось П. некоторых растений, усваивающих свободный азот воздуха.
Усвоение свободного азота совершается некоторыми бактериями, а при их посредстве и высшими растениями из сем. бобовых. Относительно последних давно было известно, что они как-то благотворно влияют на почву, именно на ее азотистый запас. Было замечено, что несмотря на большое количество азота, уносимое в их урожае, почва не только не беднеет, но, видимо, обогащается им. Но откуда бобовые растения берут азот, которым они обогащают почву: это долго оставалось непонятным. На основании господствовавшего в науке учения, что свободный азот атмосферы не может служить пищею растению, обогащение почвы при культуре бобовых растений старались объяснить побочными причинами. Только в 1888 г. благодаря успехам бактериологии Гелльригелю удалось сделать блестящее открытие, что "азотособиратели" (растения из сем. бобовых) питаются свободным азотом; он показал, что заражаясь в почве особыми бактериями, эти растения приобретают способность питаться свободным азотом воздуха. При этом на корнях их появляются особые образования, так наз. клубеньки (фиг. 1, w), которых при росте в стерилизованной почве не бывает.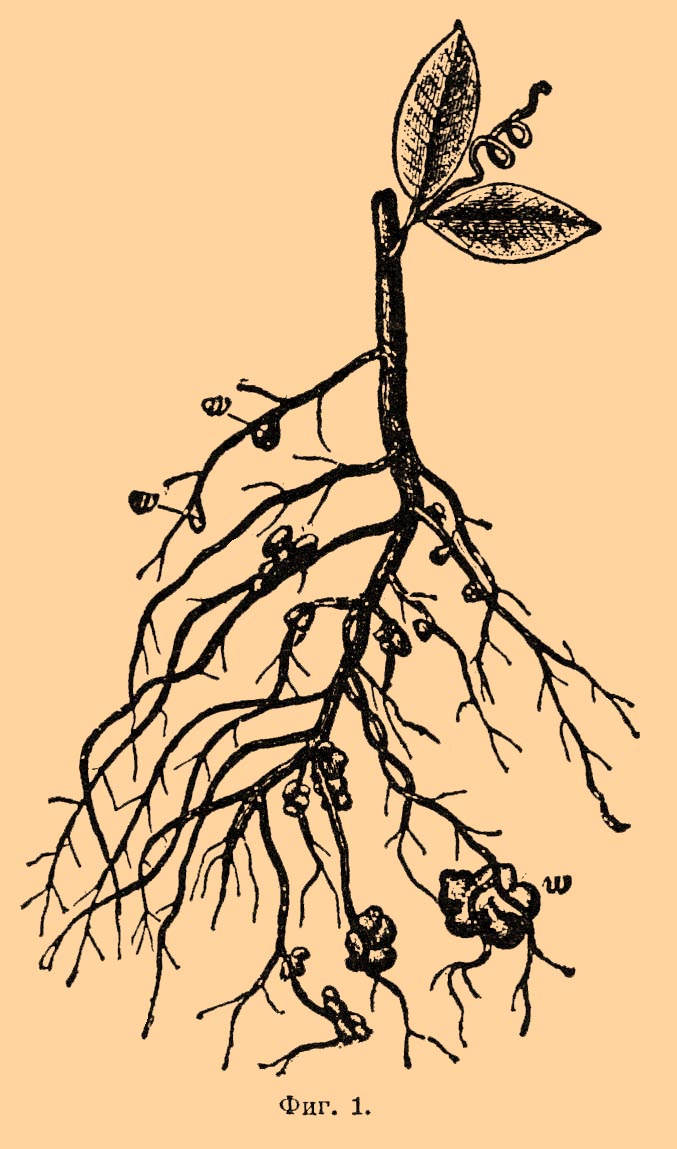
Фиг. 1.
Дальнейшие исследования, в которых приняли участие ученые всех стран, подтвердили и дополнили наблюдения Гелльригеля, так что в течение немногих лет после опубликования работы последнего вопрос этот был разъяснен с большою полнотою. Самые клубеньки на корнях бобовых растений были известны очень давно, но природа их долгое время оставалась совершенно загадочною. В 60-х годах Воронин, исследуя аналогичные образования у ольхи, высказал предположение, что у бобовых растений клубеньки происходят вследствие заражения бактериями. Открытие Гелльригеля придало особый интерес этим образованиям, и после многих споров мысль Воронина была блистательно подтверждена, главным образом Бейеринком. Бактерия Bacillus radicicola Beyer. была получена в чистой искусственной культуре; она хорошо растет в отваре бобов с сахаром при условии хорошего притока воздуха. При этом и в искусственной культуре происходит усвоение азота. Опыты заражения делались и с такими искусственными культурами путем прививки последних уколом; при этом оказалось, что образование клубеньков всякий раз происходит только в месте прививки, общего же заражения растения никогда не наблюдалось. Относительно ближайших отношений между бактерией и высшим растением кое-что остается, однако, еще неясным. Несомненно, что мы имеем здесь случай симбиоза, одинаково выгодного как для высшего растения, так и для бактерии: первое получает связанный азот, вторая — тройные соединения, которые нужны для нее в готовом виде. Но какие именно тройные соединения дает высшее растение и как пользуется оно азотом, усваиваемым бактерией, еще неизвестно. В естественных условиях заражение происходит настолько рано, что в почве, не содержащей связанного азота, растение непосредственно переходит от питания запасными азотистыми веществами семени к П. свободным азотом; но если по какой-либо причине заражение запоздает, то хотя клубеньки развиваются, но питание свободным азотом не происходит: микроб в этом случае делается паразитом. Если в почве содержатся уже азотистые соединения (азотная кислота), то образование клубеньков и усвоение свободного азота также, по-видимому, понижаются. Наконец, в оторванных от корней клубеньках не удалось констатировать усвоения азота (Тимирязев). Из растений других семейств питание свободным азотом доказано для Elaeagnus, Alnus и Podocarpus, где отношения те же, только клубеньки на корнях производятся другими низшими организмами.
Из бактерий ныне известно уже несколько видов, для которых свободный азот представляет вполне пригодную пищу. Наиболее обстоятельно изучена в этом отношении открытая и исследованная Виноградским бактерия Clostridium Pasteurianum, которая живет в почве и, по-видимому, пользуется очень широким распространением. Clostridium — анаэробный организм, т. е. может развиваться только в бескислородной среде. Но в почве он живет в симбиозе с двумя другими бактериями, образуя вместе с ними слизистые комки, напоминающие кефирные зерна. Эти спутники сами не способны усваивать свободный азот, но они поглощают кислород и тем защищают клостридий от вредного влияния этого газа. В симбиозе с ними Clostridium Pasteurianum может развиваться во воякой почве, какова бы ни была степень ее аэрации, и обогащает ее связанным азотом. Микроб производит при этом брожение сахара и на каждые 1000 частей его усваивает 1,5—3 части азота. Так как почвы относительно богаты углеродистыми соединениями и бедны азотом, то деятельность этого микроба представляется в высокой степени плодотворною.
Свободный ли азот или какое-либо простейшее соединение его воспринимается растением в качестве азотистой пищи, продуктом усвоения являются белковые вещества, синтез которых непрерывно совершается в растении. Химических превращений, при этом происходящих, не удалось еще разъяснить вполне удовлетворительно. Весьма вероятным представляется предположение, что промежуточным образованием между белковыми веществами и указанными выше простейшими соединениями азота являются некоторые амидокислоты ("амиды"), весьма распространенные в растениях, главным образом аспарагин COOH—CH(NH 2)—CH2—CO—NH2. Вещество это является и продуктом распада белков в живой клетке растения, а в присутствии углеводов снова дает белки (регенерация белков см. ниже); можно думать поэтому, что синтез этих последних проходит через ту же стадию. Из аспарагина и углеводов низшие организмы легко образуют белки своей плазмы; то же доказано и для высших зеленых растений (Hansteen). Эта гипотеза подтверждается, отчасти, и новейшими опытами синтеза белковых тел in vitro: уплотнением амидокислот получены вещества, по многим свойствам напоминающие белки (Lilienfeld), При прорастании ячменя в дистиллированной воде образуется путем распада белков вообще мало амидов; если же прорастание происходит в растворе азотнокислых или, еще лучше, аммиачных солей, где возможен, следовательно, синтез белковых тел, то количество амидокислот оказывается резко повышенным. Но, вообще, этот вопрос, конечно, еще далек от окончательного решения. Возможно, что синтез белковых тел протекает несколько различно, смотря по тому, какое вещество служит источником азота для растения. Далее, принимая во внимание, что для образования белков из азотной кислоты или аммиака нужны еще углеродистые соединения, а эти последние образуются в листьях, куда притекает с почвенным раствором и азотная кислота, нужно думать, что белковые вещества образуются зеленым растением главным образом в листьях. В пользу этого говорят и некоторые прямые наблюдения. Однако непосредственно оба синтетические процесса — образование белков и образование тройных соединений — между собою не связаны и в зеленом растении, не говоря уже о растениях, лишенных хлорофилла, у которых происходит лишь синтез белковых тел.
3) Усвоение элементов золы. Как для синтеза питательных веществ, так и для питания в тесном смысле слова растению, кроме 4 органогенов (углерода, водорода, кислорода и азота), нужны еще некоторые другие элементы. Это явствует уже из того, что каждое растение после сожигания оставляет более или менее значительное количество золы (1—10%, иногда значительно более). Однако если бы мы вздумали по химическому составу золы различных растений судить о том, какие именно элементы нужны еще растению, то едва ли пришли бы к определенному заключению. Прежде всего, как количественный, так и качественный состав золы очень неодинаков у различных растений; до сих пор в ней найдены следующие элементы: сера, фосфор, хлор, бром, йод, фтор, бор, кремний, литий, натрий, калий, рубидий, цезий, кальций, магний, стронций, барий, железо, кобальт, никель, марганец, цинк, ртуть, медь, серебро, алюминий, таллий, титан, олово, свинец, мышьяк и селен. Большинство этих элементов встречается, правда, лишь у немногих растений, но зато в некоторых случаях содержание их достигает очень значительной величины. С другой стороны, и постоянное нахождение какого-либо элемента в золе не указывает еще на необходимость его для растения, так как это может произойти и просто оттого, что этот элемент всегда находится в почвенном растворе. Вопрос о том, какие именно элементы золы необходимы для П. растений, был решен синтетически, путем искусственных культур. Но еще прежде ученым пришлось заняться вопросом, откуда растение берет вещества, находимые в золе, так как господствовавшее в свое время воззрение на сущность жизненного процесса принимало, что жизненная сила, действующая в растении, может создать все химические элементы, необходимые для последнего, из воды и воздуха. Это воззрение вполне исчезло лишь в тридцатых годах нынешнего столетия, когда наконец было признано, что и в растении химические элементы не превращаются друг в друга и что составные части золы воспринимаются растением извне, из почвы. В 1833 г. Вигманн и Польстдорф, выращивая растение с особыми предосторожностями на кусочках платиновой проволоки, смоченных дистиллированною водою, нашли, что выросшее в таких условиях растение содержит ровно столько же золы, сколько было ее в семени. Только теперь можно было заняться указанным выше вопросом, для решения которого воспользовались искусственными, так наз. водными, культурами растений, методика которых была превосходно разработана в 60-х годах, главным образом Кнопом. Прием этот и ныне весьма часто применяется для решения различных как теоретических, так и чисто практических вопросов П. растений и состоит в следующем. Семя проращивают в древесных опилках, смоченных водою, или на канве над водою до тех пор, пока не получится проросток достаточной величины. Тогда его переносят в банку подходящей формы, вроде изображенной на фиг. 2.
Фиг. 2.
В банку наливают раствор того или другого состава, о чем ниже, и закрывают пробкою, пропитанную парафином (чтобы не плесневела). В центре пробки сделано отверстие, в которое и вставляют растение при помощи ваты так, чтобы корни его были погружены в раствор, а листики находились снаружи над пробкою. Банку обертывают непрозрачною бумагою — иначе в растворе разведутся водоросли — и выставляют на свет, избегая, однако, прямого солнца. Если раствор солей составлен согласно с потребностями растения, последнее будет развиваться вполне нормально. В то же время этот метод дает легкую возможность изучать все стороны П. растений, в особенности же его потребности в пище. Вместо столь сложного субстрата, как почва, здесь — совершенно определенный раствор солей, которые мы легко можем учитывать по приемам аналитической химии, а ход развития всегда будет служить при этом для нас указателем. Путем таких опытов и было выяснено, что из почвы растению нужны лишь семь элементов: калий, кальций, магний, железо, фосфор, азот, сера; три последние в виде окислов (кислот) P 2O5, N2O5, SO3; другие степени окисления непригодны для высших растений. Их доставляют растению в следующей комбинации:
Азотно-кальциевой соли Са(NО 3)2 — 4 части
Азотно-калиевой " KNO 3 — 1 "
Фосфорно-калиевой " КН 2 РО 4 — 1 "
Серно-магниевой " MgSO 4 — 1 "
Воды — 3000—8000 ч.
Фосфорнокисл. (или хлорист.) железа — следы.
Это — раствор солей, предложенный Кнопом (существуют и другие рецепты); он очень пригоден для П. растений и всасывается ими целиком. Общая концентрация смеси не должна превышать 5 на 1000; всего лучше 1—2:1000. Ко времени цветения концентрация должна быть понижена, а во время созревания плодов для растения обыкновенно достаточно уже одной воды, так как опыты показали, что оно воспринимает большую часть минеральных веществ еще до цветения или плодоношения. Из других элементов, находимых в золе, хлор в небольшом количестве оказывается полезным в растворе, но, по-видимому, только потому, что в присутствии хлористых металлов легче сохраняется кислая реакция раствора, необходимая для успешного роста (при щелочной реакции растение страдает). Большое количество хлористого натрия в растворе, как и в почве, вредно для растения. Только солончаковые растения могут переносить значительные количества этого вещества, но и их можно выращивать и без NaCl, несмотря на привычку к соленым почвам. Кремнекислота встречается в золе почти всех растений, а в некоторых случаях содержание ее достигает значительного процента (в золе злаков до 70%, в золе хвощей 70—90%). Несомненно, что, по крайней мере, в последнем случае, вещество это полезно для растения, придавая ему значительную прочность, но собственно для П. кремнекислота, как показывают опыты водных культур злаков, не нужна растению. Присутствие всех остальных элементов, находимых в золе различных растений, объясняется следующим образом. В растение через корни поступает все то, что находится в почвенном растворе или может раствориться под влиянием кислых выделений корня. Для этого нужно только, чтобы вещество было способно диосмировать через целлюлозные оболочки и постеночный слой плазмы корневых волосков. Но если вещество проникло внутрь клеток растения, дальнейшее поступление его будет зависть от того, каким изменениям оно подвергается в них. Если вещество не нужно для П. и потому остается в растении без всяких изменений, то оно поступает лишь до тех пор, пока не установится осмотическое равновесие между почвенным раствором и клеточным соком; в этом случае зола растения будет содержать лишь незначительные количества этого вещества. Если же впитанное корневыми волосками вещество, хотя и не нужно для П., но по той или другой причине переходит в осадок (как кремнекислота, инкрустирующая клеточные оболочки) или, вообще, делается неспособным к диосмосу (напр., вследствие потребления растворителя), то поступление его в растение, согласно закону физики, будет продолжаться и зола растения будет содержать значительные количества его. Этих соображений достаточно, чтобы объяснить как присутствие в золе почти всех растений натрия, кремнезема, так и накопление более редких элементов в золе некоторых видов растений. Возможно, однако, что в этом последнем случае вещество имеет какое-либо полезное значение в жизни растения; точно это не известно. Из числа необходимых для П. зольных элементов сера нужна уже для синтеза белков, так как входит в химический состав большинства их; то же можно сказать и относительно фосфора, который входит в состав нуклеинов — главной составной части клеточного ядра, и лецитина, играющего, по-видимому, важную роль в химизме клетки. Весьма вероятно, что этим не исчерпывается роль названных элементов, но другого назначения их в настоящее время нельзя указать. Относительно железа известно, что без него не образуется хлорофилл (см. Хлороз); но, с другой стороны, по некоторым новейшим опытам, железо нужно и для грибов, не образующих хлорофилл; стало быть, и этот элемент, потребляемый растением в ничтожном количестве, играет, вероятно, не единичную роль в питании. Для чего нужны калий, кальций и магний, еще неизвестно, и нужно думать, что полный ответ на этот вопрос будет получен только после того, как мы детально узнаем все стадии внутриклеточного метаморфоза веществ. А priori ясно, что роль этих элементов должна быть очень разнообразна и что они должны принимать участие в самых различных фазах превращения веществ в клетке: они служат проводниками азотной, фосфорной и серной кислот из почвы, нейтрализуют органические кислоты и, растворяясь в клеточном соке, в высокой степени содействуют возникновению внутриклеточного давления (тургора), столь важного для роста клеток; содействуют растворению и передвижению белковых веществ, вероятно, соединяются с ними, как и с углеводами, при их химических превращениях и т. д. С этой точки зрения понятно, что указанные элементы не могут быть замещены нацело другими, тогда как частичное замещение, конечно, возможно (напр., калия — натрием, кальция — магнием), что действительно и происходит в растениях. — П. незеленых растений и в отношении минеральной пищи несколько отклоняется от П. зеленых. Кальций для грибов и бактерий не нужен, их потребности в этом отношении ограничиваются калием, магнием, железом, серой и фосфором. Два последнх элемента усваиваются некоторыми микробами и в низших степенях окисления, напр., сера в виде серноватистой кислоты и даже в виде сероводорода (бактериями серных ключей). Вполне достаточную минеральную пищу представляет поэтому для грибов и бактерий следующий весьма простой раствор (Laurent): фосфорнокислого калия KH 2PO4 — 0,75 в. ч., сернокислого магния MgSO 4 х7H 2 O — 0,10, воды — 1000 или 2000. Железа, вообще, требуется очень мало, а для этих ничтожных по величине организмов о прибавлении его нужно позаботиться только при особой чистоте всех веществ, применяемых для П. Если к этому раствору прибавить какое-либо тройное соединение и один из указанных выше источников азота, то он будет содержать все, что нужно для П. этих организмов. Более сложен состав раствора, предложенного Ролэном: сахару 70 г, винной кислоты 4 г, азотнокислого аммиака 4 г, фосфорнокислого аммиака 0,6 г, углекалиевой соли 0,6, углемагниевой 0,4, сернокислого аммиака 0,25, сернокислого цинка, сернокислого железа и кремнекислого калия по 0,07 на 1500 г воды. Aspergillus niger в этом растворе растет замечательно хорошо: сахар и аммиак под влиянием посеянной споры как бы превращаются в живое вещество грибка; каждые 3 г сахара дают один грамм сухого вещества грибка. В жидкости Ролэна обращает внимание присутствие цинка и кремния, притом в столь ничтожных дозах; для всех других растений эти элементы оказались ненужными, между тем Ролэн показал, что если исключить, напр., цинк, то рост грибка происходит втрое хуже. Новейшие наблюдения дают весьма любопытное объяснение этому. По-видимому, вещества эти действуют, как раздражители, возбуждая энергию роста микроба. Таких фактов в настоящее время известно уже немало: вещество, ядовитое (убивающее) при определенной концентрации, при более слабой — только задерживает рост, а в очень слабых дозах (в сильно разбавленном растворе) действует как раз обратно — повышает все жизненные процессы. Так, напр., при опытах с дрожжевым грибком было найдено, что развитие этого микроба:
| Задерживается при концентрации | Усиливается при концентрации | |
| Сулемою | 0,005% | 0,0003% |
| Медным купоросом | 0,025% | 0,00016% |
| Салициловою кислотою | 0,1% | 0,016% |
| Хинином | 0,25% | 0,0012% |
Явление это настолько обще, что некоторые авторы (Hueppe, Schulz, Arudt) считают его общим биологическим законом.
II. Под именем П. в тесном смысле слова подразумевают те химические превращения веществ, общие растениям и животным, которые состоят: 1) в образовании из питательных веществ (белков, тройных соединений) живой плазмы клеток и др. составных частей ее — прогрессивный метаморфоз; 2) в распадении этих последних на простейшие химические соединения — регрессивный метаморфоз веществ. Изучение сложной цепи химических превращений, при этом происходящих, и составляет главнейшую задачу физиологии, так как, с одной стороны, образование живого вещества из мертвого (инертного) органического составляет величайшую тайну природы, а с другой — изучение жизненных явлений показывает, что в основе всех их (дыхания, движения, ощущения) лежит распадение живого вещества на более простые химические соединения. Если мы вспомним, что в клетках растения параллельно с прогрессивным и регрессивным метаморфозом веществ происходят в большинстве случаев и те или другие из рассмотренных выше процессов синтеза питательных веществ и что все эти химические превращения, взаимно влияя друг на друга, представляют не ряды параллельных, но друг от друга независимых превращений, а именно одну сложную цепь или сеть их, то мы поймем, что трудность изучения предмета может равняться только интересу, им представляемому. До сих пор в этом отношении удалось выяснить лишь очень немного. Наиболее удобными для изучения представляются те случаи, где отсутствуют процессы синтеза, вследствие чего химизм несколько упрощается. Такой случай представляет, например, прорастание семян. В семени (см.) для молодого растения запасены все необходимые питательные вещества, и в первое время жизни оно питается, подобно животному, готовою пищею. Микроскопическое исследование, напр., семядолей гороха, показывает, что до прорастания клетки их набиты запасами — крахмалом и белками; во время прорастания параллельно с ростом молодого растения клетки постепенно опоражниваются и под конец бывают наполнены лишь водянистым соком, после чего засыхают и отпадают. Растворение запасов совершается при посредстве энзимов, или ферментов (см.), но может происходить и под непосредственным воздействием плазмы, что понятно уже с той точки зрения, что самые энзимы при ближайшем знакомстве с ними представляются как бы ближайшими дериватами плазмы ("осколки плазмы"). Только для жиров в растениях до сих пор с точностью не известно еще особого энзима. В акте растворения запасов нельзя не видеть аналогии с пищеварением у животных, только у растений процесс этот гораздо более прост, так как питательные вещества отложены в столь удобной форме, что не нуждаются в сколько-нибудь значительной предварительной переработке. Перейдя в раствор, запасные вещества могут уже служить для П. Некоторое представление о характере химических превращений, при этом происходящих, дают уже сравнительные химические анализы семян и проростков:
| 46 семян пшеницы | 46 проростков | Разность | |
| Вес сухого вещества | 1,665 | 0,722 | —0,953 |
| Углерода | 0,758 | 0,293 | —0,465 |
| Водорода | 0,095 | 0,043 | —0,052 |
| Кислорода | 0,718 | 0,282 | —0,436 |
| Азота | 0,057 | 0,057 | 0,0 |
| Золы | 0,038 | 0,038 | 0,0 |
Несмотря на значительное увеличение объема при прорастании, количество сухого вещества в проростке оказывается вдвое меньше, чем в семени. П. в тесном смысле сопровождается, следовательно, потерею вещества. Это происходит оттого, что и растение, подобно животному, дышит, и акт дыхания составляет для него такое же необходимое условие жизни. Процессы синтеза питательных веществ могут останавливаться (напр., разложение углекислоты — ночью) или вовсе отсутствовать: жизненный акт будет идти своим чередом; но с прекращением дыхания тотчас останавливается и жизнь. Стало быть, образование живого вещества из инертного органического сопровождается большею тратою последнего. Эти траты падают, как показывает таблица, на углерод, водород и кислород, так как при дыхании выделяются углекислота и вода. Количество азота остается неизменным; не изменяется и количество золы. Это не значит, однако, чтобы эти последние вещества не подвергались никаким химическим превращениям, но для обнаружения последних нужен уже более детальный химический анализ.
| 100 г семян Vicia sativa содержат: | Проростки, выросшие из 100 г семян того же растения (вики), содержали (через 20 дней) | |
| Белковых веществ | 28,50 | 10,60 |
| Аспарагина и амидокисл. | 2,84 | 18,05 |
| Крахмала | 37,82 | 9,93 |
| Растворимых углеводов | 5,59 | 7,67 |
| Веществ клеточной оболочки | 11,34 | 14,95 |
| Потеря в весе | 24,26 |
Из таблицы видно прежде всего сильное уменьшение количества крахмала, с избытком покрывающее потерю в весе сухого вещества. Тот же крахмал послужил, вероятно, и материалом для образования новых клеточных оболочек в проростке, так как количество клетчатки увеличилось с 11,34 до 14,95%. Путем каких именно превращений происходит этот последний процесс — еще не известно. Полагают, что углеводы не прямо переходят в вещество клеточных оболочек, а через белки, так что оболочка является продуктом превращения (окисления) белковых тел плазмы (Палладин). Далее, несмотря на указанное выше постоянство в процентном содержании а
| "БРОКГАУЗ И ЕФРОН" >> "П" >> "ПИ" >> "ПИТ" |
Статья про "Питание растений" в словаре Брокгауза и Ефрона была прочитана 6149 раз
| Коптим скумбрию в коробке |
| Крабы в кокосовом молоке |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел