
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Передвижение органов растений
Определение "Передвижение органов растений" в словаре Брокгауза и Ефрона
Передвижение органов растений
Передвижение органов растений*
— у высших растений локомоции нет; движения их состоят в перемещении отдельных органов и чаще всего происходят настолько медленно, что при поверхностном наблюдении жизни растений могут остаться незамеченными. Известно, однако, немало примеров, где П. органов происходит очень быстро, вполне напоминая соответствующие акты у животных. Различают: 1) нутационные П., происходящие лишь на молодых, еще нарастающих органах растений. Таковы изгибы стеблей и корней под влиянием силы тяжести или одностороннего освещения, завивание стеблей вокруг подпорки у вьющихся растений и проч. Все П. этого рода представляют лишь частные случаи роста (см. Гелиотропизм, Геотропизм и Рост растений); 2) вариационные П., происходящие на вполне выросших органах. Примером последних П. могут служить движения, совершаемые некоторыми растениями для ловли насекомых (см. Насекомоядные растения). У Dionaea muscipula пластинка (фиг. 1) листьев по краям усажена длинными зубцами или шипами, а по обе стороны от главной жилки на каждой половине листа находится по три симметрически расположенных щетинки, представляющих чувствительные органы листа — достаточно самого легкого прикосновения к ним, чтобы обе половинки листа моментально захлопнулись.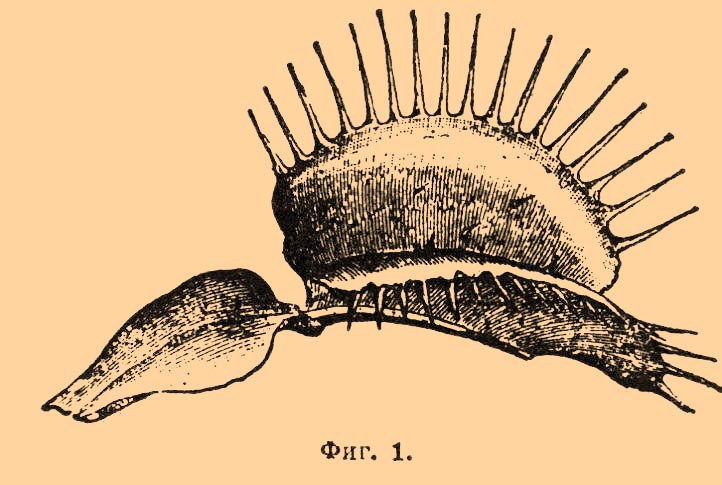
Фиг. 1.
При хорошем состоянии растения это П. происходит и в том случае, если слегка потереть верхнюю поверхность листа, не касаясь чувствительных щетинок. У Drosera rotundifolia (фиг. 2) листовые пластинки усажены красными железками; каждая железка состоит из двух частей: головки и ножки. Чувствительна к раздражению лишь головка.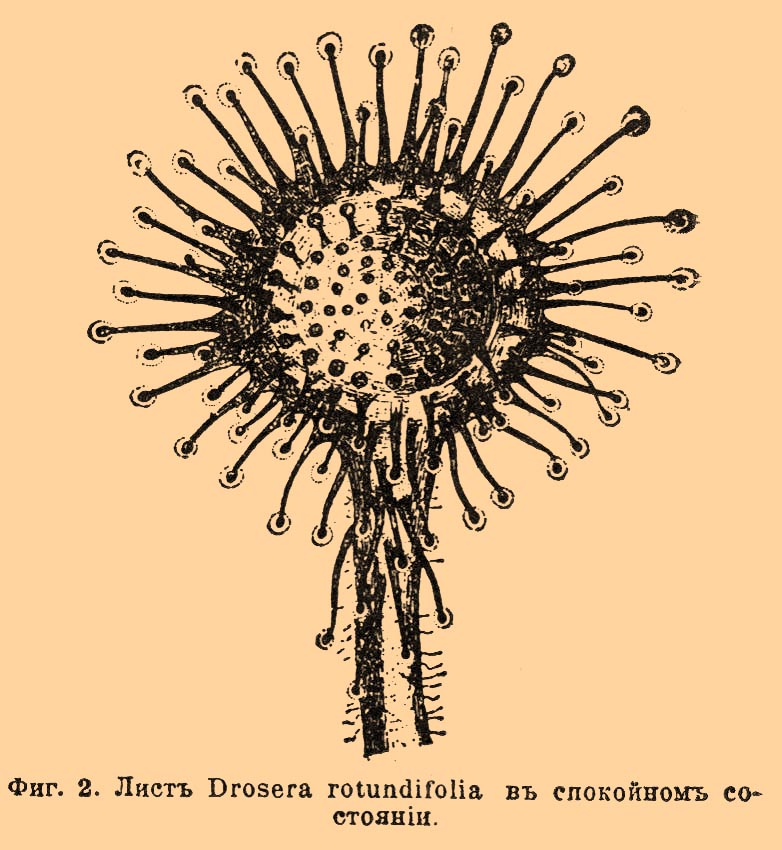
Фиг. 2. Лист Drosera rotundifolia в спокойном состоянии.
При повторном прикосновении или легком трении (в опытах Дарвина достаточно было трения, производимого кусочком волоска, весившим 0,000822 мг) уже через 10—20" железки приходят в движение, изгибаясь к центру листа; через 10—20' изгиб уже полный, и головка краевой железки прикладывается к середине листовой пластинки.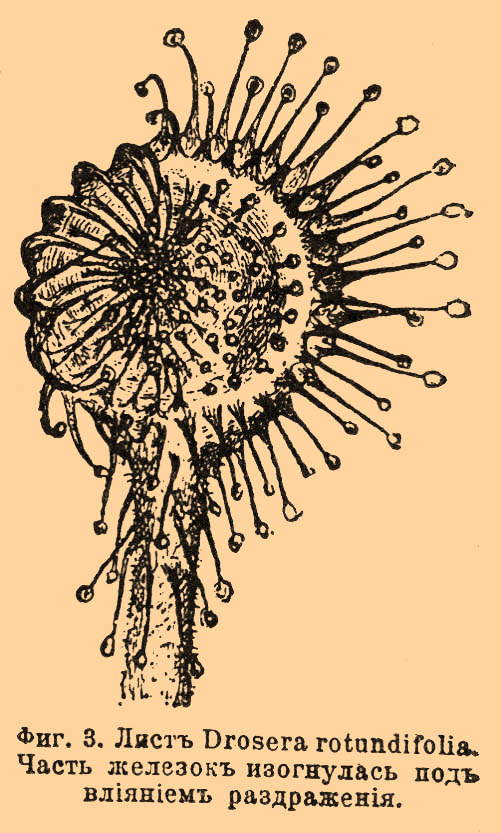
Фиг. 3. Лист Drosera rotundifolia. Часть железок изогнулась под влиянием раздражения.
Если раздражение было нанесено местно, то изгибаются прежде всего железки, непосредственно подвергшиеся раздражению, но вслед за тем раздражение распространяется по всему листу. Кроме механического раздражения, П. у насекомоядных растений вызываются и химическими раздражителями: растворами белков, аммиачных солей и др., капля раствора фосфорнокислого аммиака, содержащая 0,000423 мг этого вещества, наносимая на головку железки у Drosera, вызывает, по опытам Дарвина, уже заметное П. железки. Капли чистой воды (дождь) не оказывают действия. Известная Mimosa pudica — растение, весьма распространенное в Бразилии. Способностью к П. обладают также листья. Листья (фиг. 4) двоякоперистосложные, соединены со стеблем при помощи особого сочленения; это сочленение (а именно нижняя половина его) представляет наиболее чувствительное место у растения; такие же сочленения имеются и при соединении всех отдельных частей листа.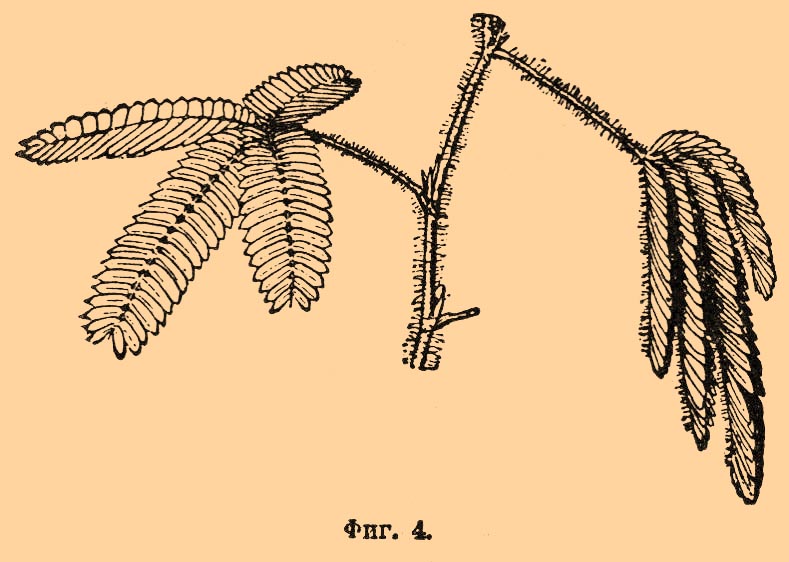
Фиг. 4.
Днем, в спокойном состоянии, главный черешок направлен косо вверх; вторичные черешки и листочки расправлены в горизонтальной плоскости. Достаточно прикоснуться к листу или, еще лучше, обжечь один листочек так называемым зажигательным стеклом, и растение тотчас придет в движение: листочки последовательно сложатся черепицеобразно на вторичных черешках; эти последние несколько опустятся и продвинутся вперед; главный черешок также опустится, и весь лист примет положение, изображенное на фиг. 4 справа. Если растение находится в хорошем состоянии, движение не ограничивается одним листом; раздражение распространяется (со скоростью 2—15 мм в 1") вверх и вниз по растению, и все листья складываются точно таким же образом. Есть указания на то, что благодаря этой способности мимоза отлично защищена от повреждения животными: привлеченные свежей зеленью, они приближаются к зарослям мимоз, но после первого же прикосновения видят перед собой лишь голые ветви и отходят прочь. Оставленное в покое, растение скоро принимает прежний вид и снова делается способным отвечать на раздражение. Если же раздражения (например, ударом) повторяются часто и непрерывно, то через некоторое время мимоза поднимает листья и делается неспособной реагировать на раздражение: чувствительность ее притупляется; после некоторого покоя она, однако, снова возвращается. Эфир, хлороформ и проч. анестезируют мимозу так же, как и животных. Чувствительностью к раздражению и способностью реагировать на него П. листьев отличаются и другие виды мимоз, а также и представители других родов и семейств, например Oxalis sensitiva и др. Только у этих растений нужно уже более энергичное раздражение, и самые П. происходят менее быстро. Аналогичные П., вызываемые внешними раздражениями, происходят в цветах многих растений; например, у Berberis чувствительными являются тычинки, которые изгибаются при прикосновении к внутренней поверхности тычиночных нитей. У других растений, например Mimulus и других Scrophulariaceae, y Bignoniaceae, П. совершаются лопастями рыльца: расправленные в спокойном состоянии, они быстро захлопываются при раздражении. У растений, снабженных усиками, легко наблюдать П. этих последних. Молодой усик находится в непрерывном движении, описывая своим кончиком круги или эллипсы различной величины; это — нутационное движение, зависящее от неравномерного роста. Встретив на своем пути подпорку, усик обвивается вокруг нее, причем контакт действует в данном случае как раздражение (подробнее см. Усики у растений).
Механизм указанных П. относительно еще очень мало выяснен. Морфологически обособленных органов или тканей для восприятия раздражения, проведения его и проявления реакции в растениях нет или почти нет. В этом отношении мы находим лишь намеки или незначительные попытки. [Факты этого рода собраны в книге Габерланда "Physiologische Pflanzenanatomie" (2 изд., 1896, стр. 473 и след.).] Упомянутые выше сочленения в листях мимозы представляют одно из наиболее ясных анатомических приспособлений для акта движения, а между тем все дело заключается лишь в том, что механические и проводящие ткани отброшены здесь к центру, чтобы не мешать изгибу, а снаружи сочленение одето толстым слоем коровой паренхимы, состоящей из живых клеточек с тонкими оболочками. И с этими ничтожными средствами растение успевает достигнуть весьма значительных результатов. Спокойное положение листа у мимозы обусловливается тем, что обе половины коровой паренхимы сочленения (верхняя и нижняя) напряжены одинаково, но действуют прямо противоположно: верхняя стремится нагнуть лист вниз, нижняя — поднять вверх. Вследствие раздражения напряжение нижней половины ослабевает, она выделяет часть воды в межклетники, уменьшается в объеме, делается дряблой, как бы вянет: лист опускается. Во время покоя, следующего за раздражением, клетки нижней половины сочленения снова насасываются водой, напряжение ткани достигает прежней величины — лист поднимается. То же изменение взаимного напряжения тканей лежит в основании и всех др. описанных выше П., только анатомические приспособления там еще менее заметны. Почему под влиянием раздражения клетки начинают выделять воду, которую только что перед этим они жадно всосали, — в этом и лежит ключ к пониманию описанных явлений. Пока мы можем только сказать, что это явление есть последствие тех физических или химических изменений, которые происходят в плазме под влиянием раздражения. Сущность этих изменений остается еще совершенно загадочной. Раздражение, как мы видели, может передаваться на значительные расстояния. Габерланд ("Das reizleitende Gewebesystem der Sinnpflanze", 1890) находит у мимозы специальную ткань, предназначенную для проведения раздражения и состоящую из живых клеток, очень похожих на ситовидные трубки; но в то же время согласно с общепринятым взглядом принимает, что передача раздражения происходит без участия плазмы, путем чисто физического (гидродинамического) процесса; к этому заключению приводит опыт, показывающий, что более сильное раздражение передается и через такие участки черешка, в которых предварительно были убиты все живые клетки. Если это и так, то несомненно, что такой способ передачи раздражения составляет особенность мимозы, между тем как во всех других случаях мы должны принять, что в процессе передачи раздражения непосредственно участвует плазма. Особой морфологической дифференцировки в клетках, передающих раздражение, мы не находим, но в настоящее время доказано, что протопласты отдельных клеток не замурованы в своих оболочках, но соединены друг с другом тончайшими нитями плазмы. Нужно думать, что именно по этим нитям в форме неизвестного еще нам молекулярного движения и передается раздражение от одной клетки к другой.
В указанных примерах П. органов происходит всегда лишь как реакция на внешнее раздражение; это — индуцированные, или паратонические, П. Если связь с внешними воздействиями более отдаленная и П. представляются нам самостоятельными, как бы произвольными, их называют автономными. Хороших примеров автономных П. взрослых органов у растений пока известно лишь очень немного; таковы П. листьев у Desmodium gyrans и губки (labellum), в околоцветнике Megaclinium falcatum (сем. Orchideae). Тройчатые листья десмодия состоят из большого верхушечного листочка и двух маленьких боковых; если растение находится в благоприятных для произрастания условиях, эти маленькие листочки совершают непрерывные П.: в то время как один опускается вниз, другой поднимается вверх, и обратно. П. происходят скачками, очень быстро (полный оборот в 1—2) и вполне независимо от каких-либо специальных раздражений, например ночью так же, как и днем. Подобные же непрерывные колебания совершает и labellum упомянутого орхидного. Промежуточный характер между автономными и индуцированными носят П., которые были известны еще в древнем мире (Плиний), а со времен Линнея получили поэтическое, но малоподходящее название сна растений (иначе никтитропные П.). Известно, что очень многие растения с заходом солнца, а иногда и раньше, особым образом складывают свои листья и закрывают цветы, так что ночное положение их резко отличается от дневного (ср. фиг. 5).
Фиг. 5. Cassia corymbosa (с фотографии). Дневное положение. Ночное положение.
У растений, листья которых соединены со стеблем при посредстве сочленений, эти П. продолжаются все время, пока лист остается жизнедеятельным, и обусловливаются изменением напряжения тканей в сочленении. При этом у видов клевера и др. листочки складываются вверх, у фасоли, лупина, белой акации и др. — вниз; у Mimosa и подобных чувствительных к раздражению растений ночное положение листьев сходно с положением их после раздражения, но при переходе в ночное положение напряжение тканей в сочленении вообще возрастает, только в верхней половине (главного) сочленения это увеличение сильнее, чем в нижней. Механизм никтитропных П., следовательно, резко отличается от П. при раздражении. Соответственно этому хлороформ, эфир не останавливают никтитропных П., тогда как чувствительность мимозы к раздражению исчезает при анестезии. У растений, не имеющих сочленений в листьях, П. последних из дневного положения в ночное и обратно происходят лишь на молодых, еще нарастающих листьях и прекращаются с окончанием роста их. Они обуславливаются преобладанием роста то верхней, то нижней поверхности листа: в первом случае лист будет опускаться, во втором — подниматься. Биологическое значение никтитропных П. видят в приспособлении к защите от ночного охлаждения путем излучения теплоты. Заметим, однако, что у некоторых растений никтитропные П. вызываются также слишком сильной инсоляцией (полуденный сон растений). Некоторые бразильские и мексиканские мимозы регулярно утром, с усилением инсоляции, складывают свои листики, вечером же расправляют их, радикально изменив таким образом часы сна и бодрствования. Что касается зависимости от внешних условий, то паратонический характер П. этого рода сказывается например в том, что ночное положение листьев можно вызвать и днем, для этого нужно только поставить растение в темное помещение. Затемняя днем и освещая ночью, можно извратить периодичность сна. Можно, наконец, заставить растение бодрствовать в течение многих суток, если держать его днем на рассеянном свете, ночью — на достаточно сильном искусственном свете. В высоких широтах (в Норвегии под 70°) в период долгих летних дней наблюдали, что мимоза не закрывает своих листьев все время, пока солнце остается над горизонтом. Однако во всех этих случаях приходится считаться и с тем, что можно назвать привычкой растения; в темноте в первые 1 — 3 — 5 дней, несмотря на полное отсутствие света, растение продолжает правильно утром раскрывать свои листья, вечером складывать их, и только постепенно эти П. делаются все более и более слабыми и, наконец, вовсе прекращаются. То же происходит и в том случае, когда растение подвергается непрерывному освещению. Равным образом, затемняя растение днем, легко вызвать ночное положение лишь в том случае, если опыт делается в послеполуденные часы. До полудня он удается не со всеми растениями, а только с наиболее чувствительными. И в естественных условиях перемена погоды (тучи) после полудня вызывает явления "сна" у большего числа растений, чем до полудня. Это неполное совпадение реакции с раздражением в никтитропных П. объясняют накоплением предыдущих раздражений, так называемого последействия света: ритмические раздражения светом в естественных условиях смены дня и ночи как бы накапливаются в растении, и эффект их (реакция) продолжает сказываться в течение некоторого времени и при постоянных внешних условиях (ср., например, сказанное в статье о гелиотропизме растений — где явления последействия света более просты и наглядны). Закрывание околоцветника, составляющее сон цветов, представляет общеизвестное явление, которое всегда легко наблюдать хотя бы на маргаритке, одуванчике, белой кувшинке. Подобно П. листьев, не имеющих сочленений, это тоже явления роста, по крайней мере в большинстве случаев: если преобладает рост верхней (внутренней) поверхности околоцветника — цветы раскрываются; и обратно, при преобладании роста нижней (наружной) поверхности цветок закрывается. Раскрывание и закрывание цветов представляет суточную периодичность, но очень различную у различных видов растений; например, у белой кувшинки цветы бывают раскрыты с 7 часов утра до 5 часов пополудни, у льна с 5 часов утра до полудня, у Cactus grandiflora — с вечера до полуночи. "Часы флоры" Линнея представляют подбор растений, раскрывающих и закрывающих свои цветы в определенные часы суток. Однако такие часы очень неточны: в пасмурные дни некоторые цветы вовсе не раскрываются или раскрываются и закрываются значительно позднее обычного. Это объясняется тем, что никтитропные П. цветов также регулируются внешними влияниями (температурой, светом), хотя здесь зависимость от внешних условий гораздо слабее, чем в П. листьев. Экспериментально влияние внешних условий во многих случаях может быть доказано вполне убедительно. Так, например, при постоянной внешней температуре, усиливая интенсивность света, легко вызвать раскрывание закрытых цветов Tulipa, Crocus, и обратно: понижая интенсивность света, вызываем закрывание их. Еще сильнее влияние температуры: всякое повышение ее (для Crocus иногда достаточно повышения температуры на ½°) вызывает раскрывание цветка, понижение — закрывание его. Повышая температуру, можно заставить цветы Crocus раскрываться в темноте, и обратно: понижая температуру, вызываем закрывание их и на свету. Комбинируя влияние света и температуры, можно сильнее влиять на никтитропные П. цветов.
П., происходящие при раскрывании плодов и рассевании семян и спор, существенно отличаются от всех предыдущих тем, что представляют уже явления физические, а не физиологические. Соответственно этому и механизм их гораздо более прост. В огромном большинстве случаев П. этого рода обусловливаются неодинаковой гигроскопичностью различных тканей, неодинаковой степенью разбухания их при впитывании воды или вообще неодинаковой напряженностью соседних слоев тканей. Явления этого рода очень разнообразны (см. Kerner v. Marilaun, "Das Pflanzenleben" (выходит русский перевод под ред. И. П. Бородина).
Литература по этому вопросу очень обширна. Укажем главнейшие сочинения. Ч. Дарвин, "Способность растений к движению"; его же, "Насекомоядные растения"; Pfeffer, "Physiologische Untersuchungen" (1873 и 1875); его же, "Zur Kenntnis d. Contactreize" ("Untersuch. aus d. botan. Inst. zu T ü bingen", т. I); Wiesner, "Das Bewegungsverm ögen de r Pflanzen"; Баталин, "Ueber die Ursachen d. periodischen Bewegungen der Blumen- und Laubbl ä tter" ("Flora", 1873); его же, "Mechanik der Bewegungen d. insektenfressenden Pflanzen" ("Flora", 1877).
Дм. Ивановский.
| "БРОКГАУЗ И ЕФРОН" >> "П" >> "ПЕ" >> "ПЕР" >> "ПЕРЕ" |
Статья про "Передвижение органов растений" в словаре Брокгауза и Ефрона была прочитана 1282 раз
| Коптим скумбрию в коробке |
| Жаренный морской черенок |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел