
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Мышцы
Определение "Мышцы" в словаре Брокгауза и Ефрона
Мышцы
Мышцы*
мускулы (Musculi — состоящие из особой ткани [мускульной, или мышечной] — см. ниже) — органы животных, которые, обладая в высшей степени способностью сокращаться в определенном направлении, служат главными активными органами движения животных (многоклеточных, а отчасти и одноклеточных) — в противоположность пассивным органам движения, играющим лишь роль опоры, рычагов для прикрепления М. и приложения их действия. В простейшей форме М. свойственны и некоторым простейшим животным (Protozoa), особенно инфузориям и грегаринам; в эктоплазме их обособляются особые тончайшие волоконца (мионемы), обладающие способностью укорачиваться (способность сократительности свойственна вообще живой протоплазме, но там сокращения не происходят непременно в одном определенном направлении). У многоклеточных (Metazoa) для образования элементов мышечной ткани служат уже особые клеточки (мускульные клеточки, или миобласты), в протоплазме которых и развиваются тонкие веретенообразные или нитевидные мускульные волоконца, или фибриллы, служащие основным элементом мышечной ткани и соответствующие мионемам простейших. Так, клеточки эпителия многих кишечно-полостных могут сохранять свое эпителиальное поверхностное положение, но образовать в основании мышечное волокно (это так наз. эпителиально-мышечные клеточки), у других кишечно-полостных (или даже у тех же, но в других частях тела) клеточки, образующие мышечное волокно, лежат глубже, под верхним слоем эпителия. Увеличение толщины мышечного слоя (а вместе с тем и силы его) достигается у кишечно-полостных тем, что слой фибрилл образует складки; складки эти могут даже совершенно обособляться от наружного эпителия, причем получаются пучки фибрилл, заключающие внутри произведшие их клеточки. Таково же в сущности происхождение значительной части мышечных волокон представителей других типов. Образование мышечного вещества может происходить различным образом, причем протоплазма клеточки, образующей мышечное волокно (миобласта), с ее ядром может сохраняться в большей или меньшей степени: мышечные фибриллы могут развиваться на одной стороне миобласта или окружать его отчасти или со всех сторон. Отдельный миобласт может производить лишь одну фибриллу или целый пучок их. Миобласт вместе с его фибриллами носит название мышечного волокна; из последних и слагаются М. Мускульные фибриллы бывают то однородны на всем своем протяжении, то состоят из отдельных мясных частичек, причем фибрилла получает видимую под микроскопом поперечную полосатость вследствие чередования частиц, сильнее преломляющих свет, с менее преломляющими. Поперечно-полосатые фибриллы одного мышечного волокна расположены таким образом, что мышечные частицы образуют пластинки (поперечные по отношению ко всему мышечному волокну), а промежуточное вещество составляет пластинки, чередующиеся с первыми. Действуя различными реактивами на поперечно - полосатое мышечное волокно, мы можем вызвать распадение его или на волоконца (напр. действием спирта), или на поперечные пластинки (действием желудочного сока) и мясные частицы. Поперечно-полосатые мышечные волоконца встречаются уже у простейших и кишечнополостных, вообще же поперечно-полосатые волокна встречаются там, где М. производят более энергические, быстрые сокращения. У суставчатоногих не только М. органов движения, но и М. стенок кишечника поперечно-полосаты, у моллюсков все М. гладкие, у позвоночных М. скелетные и вообще произвольные, а также М. сердца поперечно-полосаты, М. большей части кишечного канала, кровеносных сосудов и других внутренностей — гладкие. Форма мышечных волокон как гладких, так и поперечно-полосатых обыкновенно веретенообразная или цилиндрическая, реже разветвленная на концах (напр. в толще тела гребневиков) или образующая сеть (напр. в М. сердца позвоночных, в кишечнике некоторых суставчатоногих). Мышечное волокно может заключать в протоплазме одно или много ядер; умножение их происходит, насколько известно, не путем слияния нескольких миобластов для образования мышечного волокна, а путем деления первоначального ядра миобласта. У позвоночных скелетные М. состоят из поперечно-полосатых волокон, в которых можно отличать 1) наружную однородную оболочку (сарколемму), 2) поперечно-полосатое волокнистое вещество и 3) многочисленные ядра с остатками протоплазмы миобласта; кроме того замечается специальное нервное окончание в виде скопления протоплазмы с ядрами под сарколеммой, в которое входит веточка двигательного нерва. Различают два рода поперечно-полосатых мышечных волокон позвоночных: более бледные с менее многочисленными ядрами и более темные с многочисленными ядрами. Сокращения первых быстрее и менее продолжительны. Оба рода волокон комбинируются в различных количествах в разных М. Отдельные мускульные волокна слагаются в пучки и одеваются соединительной тканью, образуя отдельные массы М., окруженные соединительнотканной оболочкой — фасцией. Для прикрепления их к костям служат особые плотные образования из соединительной ткани — шнуровидные сухожилия (Теndines) или плоские, перепончатые сухожильные растяжения (Aponeuroses). Роль сухожилий и сухожильных растяжений заключается как в соединении М. с костями, так и в передаче действия М. на расстояние: многие М. прикрепляются к костям, приводимым ими в движение, с помощью очень длинных сухожилий (в конечностях). М. сердца позвоночных состоят из коротких мышечных клеточек с поперечно-полосатым содержимым, слагающимся в разветвленные и соединяющиеся (анастомозирующие) между собой ряды (см. Сердце). Гладкие мускульные волокна позвоночных имеют форму веретенообразных клеточек, более или менее удлиненных и располагающихся по большей части в виде продольных и поперечных слоев. У человека и позвоночных поперечно-полосатые М. (кроме сердца) зависят от воли, гладкие не зависят. Первые, за исключением кольцевых М., или сфинктеров (Sphincter), окружающих отверстия и служащих для их замыкания, оканчиваются на обоих концах сухожилиями или сухожильными растяжениями; средняя часть, состоящая собственно из мышечной ткани, называется обыкновенно брюхом (venter) М.; между сухожилиями отличают начальные и конечные (или головку — Caput и хвост — Cauda M.). Иногда брюхо М. подразделяется сухожилием надвое, тогда М. называется двубрюшной (biventer); если притом сухожилие не в виде шнурка, а в виде перегородки с зубчатыми выростами, вдающимися в М., то такое сухожилие называется Inscriptio tendinea. Если сухожилие вдается в М. и мышечные пучки прикрепляются к нему с двух сторон под острыми углами, то М. называется перистой (pennatus), если с одной стороны — полуперистой (semipenatus). Если начальных сухожилий несколько, то М. называется соответственно числу их двуглавой (biceps), трехглавой (triceps), четырехглавой (quadriceps). По форме различают длинные М., широкие, толстые (широкие со значительной толщиной) и круговые (см. выше). Далее, М. делятся по их функции (сгибающие и разгибающие, поднимающие и опускающие, приводящие, т. е. приближающие конечность к средней линии тела, и отводящие и т. д.); если несколько М. действуют в одном смысле (напр. сгибатели данного сочленения), то их называют коадъюторами (Coadjutores); если действие их противоположно (напр. сгибатели и разгибатели), то они называются антагонистами (Antagonistae). Вследствие большой сложности мышечной системы отдельные М. не могут быть здесь рассмотрены. Представление о главнейших из них дает приложенная таблица, а также см. статьи: Брюхо (с фиг.), Грудная клетка и фиг. Грудные внутренности, Дыхание. Данные по сравнительной анатомии мышечной системы см. в статьях об отдельных типах и классах животного царства. Здесь же рассмотрен общий план строения мускульной системы позвоночных, насколько это необходимо для правильного понимания строения ее у человека. Изменения в строении мышечной системы могут происходить благодаря распадению мускула на два следующие друг за другом отдела, разделенные сухожилием, благодаря продольному расщеплению, благодаря распадению на слои, благодаря слиянию первоначально отдельных М., благодаря увеличению или уменьшению М. или полному исчезанию их (причем они заменяются часто соединительнотканной перепонкой — фасцией), наконец, благодаря изменению точек прикрепления М. Оставляя в стороне гладкие мышечные волокна, мы можем различать в мускульной системе позвоночных 1) кожную мускулатуру и 2) скелетную; в состав последней входят а) париетальная мускулатура с ее производными: мускулатурой конечностей, глазного яблока и диафрагмой и б) висцеральная мускулатура — мускулатура головы кроме глазных и некоторых других М.
I. Кожная мускулатура, слабо развитая у рыб и земноводных, значительно развита у пресмыкающихся и птиц и наибольшего развития достигает у млекопитающих (напр. у ежа, броненосцев, ехидны и др.). У человека слабый остаток ее представляет Platysma myoides (фиг. II, 9).
II. Скелетная мускулатура. а) Париетальные М. состоят в простейшей, типичной для позвоночных форме из двух мышечных масс, спинной и брюшной, с каждой стороны тела и хвоста. Эти 4 продольные массы сталкиваются на спинной и брюшной средней линии и на боковых линиях тела (лишь у ланцетника — Amphioxus, принадлежность которого к позвоночным, впрочем, спорна, боковых линий нет). Поперечные перегородки из соединительной ткани — миокоммы (Myocommata) делят продольные мышечные массы на отделы — миомеры, или сомиты (Myomeren, Somiten немецких авторов); каждому из последних соответствует позвонок и пара спинно-мозговых нервов. Этот основной план строения хорошо виден у рыб, хотя уже у них на брюшной стороне обособляются некоторые группы М. — зачатки прямой и косых брюшных М. Дифференцирование спинной половины париетальных мускулов отстает от дифференцирования брюшных во всем типе позвоночных; у рыб спинные М. сохраняют еще первоначальный характер. Процесс обособления идет дальше у двоякодышащих рыб и у земноводных; у хвостатых земноводных обособляется поперечная брюшная М. Значительно далее идет дифференцирование (и вместе с тем сглаживание первичного строения мышечной системы) у пресмыкающихся; в связи с обособлением грудной и брюшной области и париетальная мускулатура изменяется; она дает в грудной области наружные и внутренние межреберные M. (Musculi intercostales externi et interni), a в поясничной области мускулатура теряет свой сегментированный характер, образуя широкие массы, тянущиеся от задних ребер к тазовым костям (напр. Musculus quadratus lumborum по бокам позвоночного столба). В то же время в спинной половине париетальной мускулатуры дифференцируется целый ряд М. (напр. levatores costarum, scaleni, longissimus и др.). В области клоаки и основания хвоста также появляются новые М., между тем как собственно хвостовые М. сохраняют примитивный характер. У птиц первоначальный характер мускулатуры еще более затемнен, особенно вследствие громадного развития мускулатуры передних конечностей. У млекопитающих боковые брюшные М. (M. obliquus externus, internus и transversus) за исключением тупайи (Tupaja) совершенно несегментированы; прямая М. имеет различное число Myосоmmata (см. фиг. в статье Брюхо). В грудной области, косым М. брюха соответствуют и здесь межреберные М. Спинная половина париетальной мускулатуры представляет сильную дифференцировку. Мускулы глазного яблока см. в статье Глаз. Мускулатура конечностей, как показывает и строение, и история развития ее у животных с наиболее примитивным строением этих органов (рыб), может считаться за производное париетальной мускулатуры, и притом частью спинной, частью брюшной половины ее. Мускулатура плавников (парных) рыб сводится, главным образом, к М. поднимающим, опускающим и приводящим конечность. Гораздо большую сложность представляет мускулатура конечностей у всех остальных позвоночных в связи с тем, что конечность из плавника превращается в систему рычагов, представляющую орган хождения — акта несравненно более сложного, чем плавание. Сложность эта все увеличивается и достигает своей высшей степени в конечностях приматов (обезьян и человека). Другим производным париетальной мускулатуры надо считать грудобрюшную преграду (см.), или диафрагму (Diaphragma). Первые зачатки ее встречаются уже у хвостатых земноводных (Urodela) в виде вдающихся между околосердечной сумкой и печенью мышечных волокон верхнего конца поперечной брюшной М. У черепах, особенно у крокодилов и птиц она выражена резче и образована М., отходящими частью от позвоночника, частью от ребер. Полного развития она достигает лишь у млекопитающих. б) Висцеральная мускулатура состоит из М. висцерального скелета (см. Скелет), т. е. из М. жаберного аппарата и М. жевательных. У высших позвоночных с исчезновением жаберного дыхания и соответственные части мускулатуры подвергаются сильному упрощению и изменению, однако, на шее и в дне ротовой полости встречается ряд мускулов, относящихся к висцеральной мускулатуре, и именно жаберному ее отделу, частью появляющихся уже у земноводных, частью свойственных именно млекопитающим; к первым относятся напр. M. mylohyoideus, sterno-, оmо-, geniohyoideus и др., к последним stylohyoideus, styloglossus, stylopharyngeus. (Жевательные M. см. в соответственной статье). Кроме того к висцеральной мускулатуре относится мускулатура языка. Мимические М., от которых зависит мимика лица, заметные в виде незначительных зачатков уже у низших позвоночных, достигают значительного развития у млекопитающих, и притом развитие это идет параллельно с развитием головного мозга и психической жизни. М. эти, как показывает сравнительно анатомическое исследование, представляют дифференцированные участки первоначально более однородного мышечного покрова, одевавшего голову и шею и сохранившегося отчасти в недифференцированном виде, как Platysma myoides.
Н. Кн.

МЫШЦЫ ЧЕЛОВЕКА.
Фиг. 1. Вид сзади. На правой половине тела поверхностные мышцы отчасти удалены. 1. Затолычная мышца (Musculus occipitalis). 2. Треугольная мышца головы и шеи (M. splenius capitis et colli). 3. Височная мышца (M. temporalis). 4. Мышцы век, или круговые глаза (М. orbicularis s. sphincter palpebrarum). 5. Жевательная мышца (М. masseter). 6. Мышца, наклоняющая голову, или грудино-сосковая (М. sterno-cieidomastoideus). 7. Мышца капуциновая, или трапецевидная (М. cucullaris s. trapezius). 8. Мышца, поднимающая опатку (М. levator scapulae s. angularis). 9. Ромбоидальная мышца (М. deltoideus). 10. Верхняя мышца лопатки (М. supraspinatus). 11. Нижняя мышца лопатки (М. infraspinatus). 12. Большая круглая мышца (М. teres major). 13. Дельтовидная мышца (М. deltoideus). 14. Широкая мышца спины (М. latissimus dorsi). 15. Длинная спинная мышца (М. longissimus dorsi). 16. Задняя нижняя зубчатая мышца (М. serratus posticis inferior). 17. Гребень подвздошной кости. 18. Мышца трехглавая плеча (М. triceps s. extensor brachii). 19. Внутренняя локтевая мышца (М. ulnaris internus). 20. Мышца, разгибающая пальцы общая (М. extensor digitorum communis). 21. Мышца, поворачивающая лодонь кнаружи (сверху) длинная (М. s upinator longus). 22. Мышца, поворачивающая лодонь кнаружи (кверху) короткая (М. supinator brevis). 23. Четвертая мышца локтя (М. anconeus quartus). 24. Мышца, разгибающая большой палец длинная (М. extensor pollicis longus). 25. Мышца длинная, отводящая большой палец (М. abductor pillicis longus). 26. Общая тыльная связка запястья (Ligamentum carpi commune dorsale s. armillare). 27. Сухожилия мышцы, распрямляющей пальцы. 28. Межкостные мышцы (М. interossei). 29. Большая ягодичная мышца (М. glutaeus magnus). 30. Малая ягодичная мышца ((М. glutaeus mimimus). 31. Грушевидная мышца (М. pyriformis s. pyramidalis). 32. Верхняя двойничная мышца (М. gemellus superior). 33. Наружняя запирательная мышца (М. obturator s. obturatorius externux). 34. Нижняя двойничная мышца (М. gemellus inferior). 35. Квадратная бедренная мышца (М. quadratus femoris). 36. Двуглавая бедренная мышца (М. biceps fempris). 37. Полусохожильная мышца (М. semi tendinosus). 38. Полуперепончатая мышца (М. semi membranosus). 39. Двуглавая икроножная мышца (М. gemellus surae s. gastrocnemius). 40. Длинная пяточная мышца (М. plantaris). 41. Пяточная мышца (М. soleus). 42. Ахиллесово сухожилие (Tendo Achillis). 43. Короткая малоберцовая мышца (М. peroneus bravis). 44. Крестообразная связка (Ligamentum cruciatum). 45. Сухожилия, разгибающие пальцы.
Фиг. 2. Вид спереди. На левой стороне поверхностные мышцы отчасти удалены. 1. Лобная мышца (М. frontalis). 2. Височная мышца (М. temporalis). 3. Мышца, смыкающая века (М. orbicularis s. sphincter palpebrarum). 4. Мышца, сжимающая нос (М. compressor nasi). 5. Кольцевая мышца рта (М. orbicularis oris). 6. Ланитная мышца (М. bussinator s. buccalis). 7. Скуловая мышца (М. zygomaticus). 8. Мышца, опускающая нижнюю губу(М. depresspr labii inferioris s. quadratus men t i). 9. Широкая шейная мышца (Platisma myoides s. subcutaneus colli). 10. Грудино-сосковая мышца (М. sterno-cieidomastoideus). 11. Грудино-подъязычная мышца (М. sterno-hioideus). 12. Шейнореберные мышцы (М. scaleni). 13. Подключичная мышца (М. subclavius). 14. Большая грудная мышца (М. pectoralis major). 15. Малая грудная мышца (М. pectoralis minor). 16. Межреберные мышцы (М. intercostales). 17. большая передняя зубчатая (М. serratus anticus major). 18. Дельтовидная мышца (М. deltoideus). 19. Двуглавая мышца (М. biceps). 20. Мышца, поворачивающая ладонь книзу (внутрь) круглая (М. pronator rotundus s. teres). 21. Мышца, поворачивающая ладонь кнаружи длинная (М. supinator longus). 22. Мышца лучевая внутренняя (М. radialis internus). 23. Мышца, сгибающая пальцы поверхностная (М. flexor digitorum sublimis). 24. Внутренняя локтевая (М. ulnaris internus). 25. Мышца, сгибающая пальцы глубокая (М. flexor digitorum profundus). 29. Сухожилия, сгибающие пальцы. 30. Мышца ладонная короткая (М. palmaris brevis). 31. Прямая брюшная мышца (М. rectus abdominis). 32. Косая внутренняя брюшная мышца (М. obliguus abdominis internus). 33. Внутренняя поясничная мышца (М. iliacus internus). 34. Мышца, направляющая влагалище бедра (М. tensor fasciae latae). 35. Портняжная мышца (М. sartorius). 36. Длинная приводящая бедра (М. adductor longus). 37. Длинная головка разгибающей голень четырехглавой, или прямая голени (М. rectus cruris). 38. Наружная головка разгибающей голень, или широкая наружная (М. vastus externus). 39. Внутренняя головка разгибающей голень, или широкая внутренняя мышца (М. vastus internus). 40. Средняя головка разгибающей голень, или широкая средняя (М. vastus medius s. cruralis). 41. Тонкая бедренная мышца, или прямая внутренняя (М. gracilis s. sectus internus). 4 2. Сухожилие мышцы, разгибающей голень с коленной чашечкой. 43. Сухожилие портняжной мышцы. 44. Длинная малоберцовая мышца (М. peroneus longus). 45. длинная разгибающая пальцы общая (М. externum digitorum communis longus). 46. Короткая малоберцовая мышца (Мю peroneus brevis). 47. Двуглавая икроножная мышца (М. gastrocnemius). 48. Крестообразная связка (Ligamentum cruciatum). 49. Сухожилия, разгибающие пальцы.
М. суть органы активных движений животных, и в наиболее развитой форме они встречаются у млекопитающих, птиц, амфибий и рыб, и из одного только взгляда на приведенную таблицу (М. человека) легко видеть, как высоко развита мышечная система человеческого организма. Функция М., состоящая в способности производить самые разнообразные активные движения, основана на их резко выраженной сокращаемости. Это свойство является лишь усовершенствованной формой элементарной сократительности протоплазмы, которая у амеб, лейкоцитов и др. выражается в способности их производить неправильные амёбоидные движения с высовыванием и втягиванием протоплазменных отростков, причем формы, через которые проходит протоплазма, до бесконечности варьируют. М. же при сокращении проходят через серию всегда одинаковых фаз: а именно, при покое волокна ее расслаблены и вытянуты в длину, а при деятельности частицы сократительного мышечного вещества располагаются всегда так, что укорачивают и в то же время утолщают М. Такая специальная форма мышечного сокращения является последствием ее анатомического устройства (см. Анатомию М.) и ее физико-химич. свойств. Другое основное свойство М. — это ее возбудимость, или раздражительность, т. е. способность ее переходить в деятельность под влиянием разнообразных раздражителей: механических, химических, термических, электрических, световых и нервного возбуждения или нервных импульсов. При нормальных условиях все поперечно-полосатые М. скелета, подчиняющиеся нашей воле, т. е. так наз. произвольно двигательные М., приводятся в деятельность нервными импульсами или нервным возбуждением, посылаемыми к ним по двигательным нервам из нервных центров спинного или головного мозга. Исключением из этого правила являются только поперечно-полосатые М. сердца, не подчиняющиеся воле и работающие под влиянием импульсов, развиваемых местными периферическими центрами сердца; поэтому разъединение всех скелетных М. от центральной нервной системы путем перерезки связывающих их между собой нервов обрекает М. на полный паралич, т. е. на полное бездействие, несмотря на то, что они при непосредственном раздражении их электрическими или механическими раздражителями и даже химическими отвечают на них сокращением. Некоторые опыты доказывают при этом, что мышцы обладают собственной раздражительностью или возбудимостью, независимой от разветвляющихся в них нервов. Таким образом, М. присущи два основных жизненных свойства: сократительность и собственная возбудимость, или раздражительность; нервное возбуждение, или нервные импульсы, посылаемые из центральной нервной системы по нервам к М. скелета, являются только необходимым толчком к переведению потенциальных сил мышечного вещества в живые силы массового движения, теплоты и электричества; такой эффект достигается тем, что волнообразное молекулярное движение нервного возбуждения, достигая по нервам до М., вызывает в них взрывчатое разложение их нестойкого вещества с освобождением живых сил. Менее совершенная форма мышечной ткани — гладкие М., заведующие движением органов так наз. растительной жизни, как то: кишечного канала, кровеносных и лимфатических сосудов, бронхов, а также и органов мочеполовой сферы — мочеточников, матки, не подчиняются (за исключением М. мочевого пузыря) действию воли и поэтому зовутся непроизвольными М.; они приводятся в деятельность или чисто рефлекторно, или автоматически, благодаря как заложенным в них нервным узлам, так и способности гладких М. к самостоятельной, независимой от нервов деятельности. Этой автоматичностью (Энгельманн) гладкие М. всего существеннее отличаются от поперечно-полосатых М. В этом отношении гладкие М., как менее дифференцированные образования, обнаруживают одно из общих свойств сократительной протоплазмы к автоматическим сокращениям. Благодаря этому свойству гладких М., кишечное содержимое передвигается постепенно по длине кишечного канала, сосуды сокращаются или расширяются и распределяют кровь по телу, моча гонится по мочеточникам в мочевой пузырь, бронхи сокращаются и расслабляются, и т. д., и все это вполне независимо от воли, от которой главным образом зависит деятельность только скелетных М. нашего тела. О механизме мышечного сокращения судят на основании детального анализа морфологических, физических и химических изменений, претерпеваемых М. при переходе их из покоя в деятельность. Микроскопические исследования М. в покоящемся и в деятельном состоянии доказывают, по-видимому, что единственными сократительными элементами являются так наз. толстые двоякопреломляющие диски, тогда как светлое, более жидкое однопреломляющее промежуточное вещество и тонкие диски играют пассивную механическую роль (см. анатомию М.). При сокращении М. светлое промежуточное между толстыми дисками вещество почти совершенно исчезает, толстые диски сближаются почти до полного соприкосновения, отделяясь друг от друга одними тонкими дисками и тонкой полоской светлого промежуточного вещества, зато толстые диски увеличиваются в ширину, вероятно, за счет всасывания ими светлого, более жидкого промежуточного вещества.
Фиг. 1. Мышечные волокна, подвергающиеся сокращению, по Энгельману. Воолкно у а находится в покое, у б — сокращение начинается, у в — достигает максимальной силы; х — толстые диски, у — светлое промежуточное вещество с тонким диском, z — в середине. Слева — волокно в обыкновенном свете, справа в поляризованном.
Особенно ясно явления эти наблюдаются на М. насекомых (Hydrophilus, Telephorus melanurus) и в поляризованном свете. Ранвье дает следующую теорию мышечного сокращения, опираясь на микроскопические изменения М. во время деятельности: доказав предварительно, что толстые диски обладают сократительностью, он указывает, что они из формы удлиненных палочек, какую имеют при покое, стремятся при возбуждении, подобно всякой сократительной саркоде, принять форму более шаровидную. Отсюда — уменьшение мышечного волокна в длину и увеличение в толщину; но это укорочение усиливается еще тем, что выдавливаемая при сокращении толстых дисков мышечная плазма располагается по краям их, где она встречает наименьшее сопротивление, вследствие чего мышечные волокна еще более утолщаются. Что же касается промежуточного светлого вещества, то оно, по Ранвье, обнаруживает крайне легкую перемещаемость и упругость, вследствие чего всякого рода укорочения М. не только активные, но и пассивные, обусловливают его перемещение из продольного направления волокна в поперечное, т. е. в его толщину. Энгельман полагает, что при сокращении толстый анизотропический диск сохраняет свою длину, но разбухает в поперечном направлении за счет поглощения воды из светлого промежуточного вещества, и это поглощение растет пропорционально с силой сокращения до полного почти исчезания светлого промежуточного вещества. Другие взгляды на механизм мышечного сокращения представляются еще довольно спорными. М., по этому, представляют такие сократительные органы, которые в сравнительно малом объеме заключают в себе массу мельчайших сократительных частиц в виде толстых дисков. Эта раздробленность сократительного мышечного содержимого, конечно, увеличивает поверхность обмена между ним и окружающими его соками и тем способствует быстроте мышечных сокращений; сообразно с этим в М. гладких, медленно сокращающихся, не наблюдается этой раздробленности на диски, тогда как в М. насекомых, отличающихся особенной быстротой сокращений, каждый толстый диск дробится еще на пять и более частей. Сближение толстых дисков при сокращении, ведущее к тому, что вся поперечная исчерченность М. при этом становится тоньше и чаще, обнаруживается еще, как это показал Ранвье, изменением мышечного спектра. Если перед щелью миоспектроскопа Ранвье поместить поперек портняжью М. лягушки с параллельными волокнами, то при искусственном ее сокращении или волевом мышечный спектр всякий раз становится шире. При расслаблении же М. или растягивании ее получается, наоборот, сужение мышечного спектра. Для детального исследования мышечного сокращения, как механического акта, прибегают к графическому методу, т. е. к записыванию при посредстве миографов этих сокращений на движущемся закопченном листе бумаги (фиг. 2).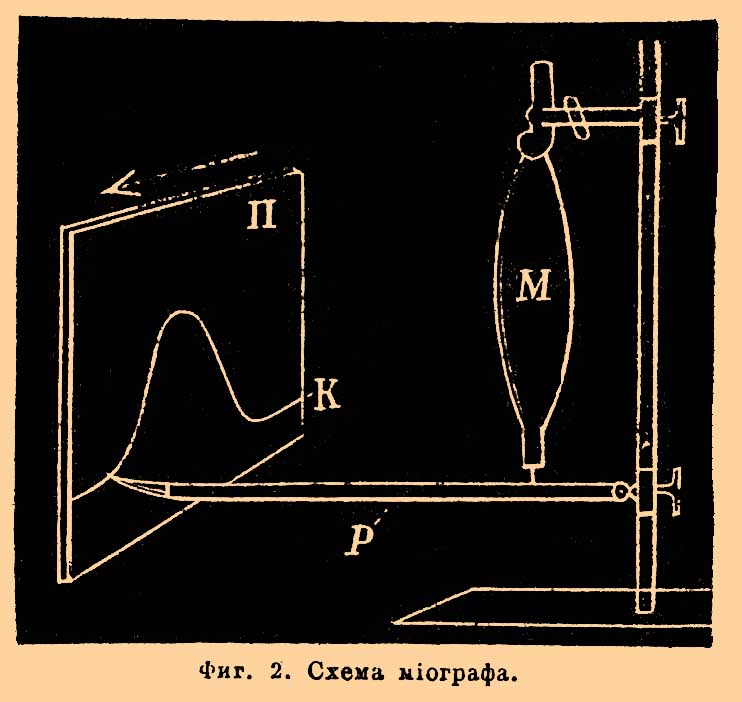
Фиг. 2. Схема миографа.
М., вырезанная из тела (или находящаяся в своем естественном месте), будучи закреплена одним своим концом, другим двигает, во время сокращения своего, рычаг Р, слегка натянутый противовесом или пружиной для вытяжения М. в момент ее расслабления. Это составляет записывающий аппарата миографа; воспринимающая же его часть состоит обыкновенно из закопченного цилиндра; равномерно вращающееся перо миографа слегка касается закопченной поверхности и при покое М. чертит прямую линию, при сокращении же дает волнообразную линию, так называемую кривую мышечного сокращения К. Чаще всего раздражают живой нервно-мышечный препарат, т. е. нерв со связанной с ним М. или индукционными ударами, или одиночными, или целой суммой их, быстро следующих друг за другом, так назыв. npepывистым током, доставляемым обыкновенно индукционным аппаратом Дю-Буа-Реймона. Включенный в первичную спираль индукционного аппарата миниатюрный электромагнит электрического сигнала Депрэ отмечает на закопченной поверхности вертящегося цилиндра момент падения на нерв или М. раздражающего электрического удара, а перо миографа — как начало, так и все последующие фазы сокращения. Волнообразная линия, записанная на той же закопченной поверхности вращающего цилиндра вибрирующим камертоном, дает в форме волн время в сотых или тысячных долях секунды (см. Хронограф), и таким образом имеются налицо все данные для графического исследования акта мышечного сокращения как в пространстве, так и во времени. Мышечные сокращения бывают двоякого рода: простое одиночное мышечное сокращение, или спазм, и тетаническое мышечное сокращение, состоящее из ряда суммированных простых мышечных сокращений. Первая форма сокращения получается при раздражении М. или связанного с ней нерва одиночным индукционным ударом, вторая же при раздражении целым рядом ударов, быстро следующих друг за другом, более 16 в сек. Кривая простого одиночного сокращения икроножной М. лягушки, изображенная на фиг. 3, указывает следующие составные фазы: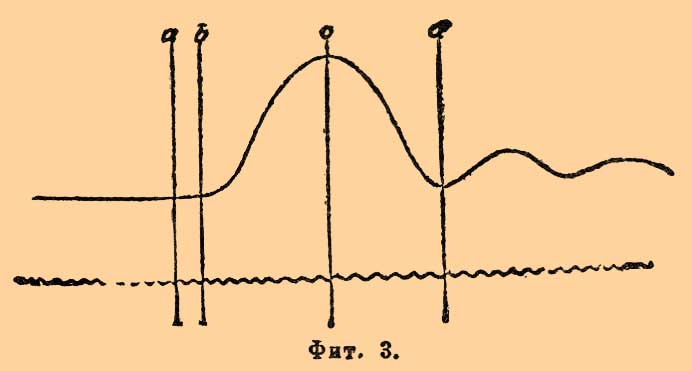
Фиг. 3.
1) период скрытого возбуждения, отвечающий промежутку от а до b, т. е. от момента падения удара на М. до начала ее сокращения, когда М., не представляя еще никаких морфологических изменений, а только молекулярные, как бы подготовляется к последующему сокращению; период этот длится обыкновенно для поперечно-полосатых М. тела около 1/100 сек.; 3) период укорочения М. вначале медленного, затем более быстрого, достигающего своего максимума у с и длящийся около 4/100 сек., и 3) период расслабления, выражающийся сначала медленным, затем более быстрым опусканием рычага, пока М. у d не достигнет своей первоначальной длины, длящийся около 5/100 сек. Весь акт отдельного мышечного сокращения пробегает, следовательно, в 1/10 сек., и это относится к случаю, когда раздражается непосредственно М.; при сокращении же, вызываемом с нерва, время это будет несколько больше, так как нервному импульсу, возникшему в той или другой точке нерва, приходится пробежать известный участок его прежде, чем дойти до М. Две меньшие волны, замечаемые после d, зависят от колебания рычага миографа. Если за отдельным индукционным ударом следует через короткий промежуток времени другой, а за ним третий, четвертый и т. д. так, чтобы предыдущее сокращение не успело еще закончиться, как посылается уже импульс к последующему сокращению, то отдельные мышечные сокращения ведут к развитию столбняка, или тетануса.
Фиг. 4.
Так, на фиг. 4, изображающей двойную мышечную кривую, первая волна сокращения только что достигла максимума своей высоты а, как наступила вторая волна сокращения b от второго индукционного удара в виде надставки на первой волне; если бы последовал быстро и третий удар, то на второй волне выросла бы еще третья надставная волна, и т. д., с той только особенностью, что высота каждого последующего сокращения бывает меньше предыдущей и это уменьшение становится тем резче, чем большее число ударов получила М. Поэтому наступает такой момент, когда быстро следующие друг за другом удары уже не вызывают дальнейшего поднимания рычага, т. е. укорочения М., а просто поддерживают существующее сокращение на определенной высоте (фиг. 5 от b до с), М. переходит в расслабление (в точке с) или вследствие истощения, или вследствие прекращения раздражения.
Фиг. 5.
На фиг. 5 от a до с зубчики суммированных отдельных мышечных сокращений, из которых складывается мышечный столбняк, не видны вследствие медленности вращения цилиндра, на котором пишет перо миографа. Все виды М. способны давать при указанных выше условиях эти две формы мышечных сокращений — простого одиночного сокращения и столбняка, или тетануса, с той только разницей, что наименее подвижными оказываются непроизвольные гладкие М. За ними идут красные поперечно-полосатые М. и наиболее быстрыми — белые поперечно-полосатые М. Гладкие М. млекопитающих требуют для совершения полного сокращения более минуты, причем на скрытый период возбуждения выпадает промежуток в 0,85 сек., т. е. почти в 100 раз больший, чем для поперечно-полосатых М.; период расслабления длиннее периода сокращения в 5 — 7 раз (Сертоли). Понятно, что при такой длительности отдельных сокращений гладких М. достаточно, чтобы на них падали раздражающие удары по разу через каждые 5 сек., чтобы дать в результате мышечный столбняк, в то время как для столбняка красных М. кролика требуется не менее 10 возбуждений, а для бледных М. по крайней мере 20 возбуждений в сек. Вообще, чем более продолжительность каждого отдельного сокращения, тем при менее частых раздражающих ударах может получаться мышечный столбняк; согласно с этим черепашьи М., отличающиеся чрезвычайной медленностью своих сокращений, дают столбняк при падении на них всего трех раздражающих ударов в 1 сек., и напротив, птичьи М., отличающиеся быстротой своих сокращений, требуют для той же цели не менее как 70 электрических ударов в секунду. Даже одни и те же M. y различных людей требуют неодинаковой частоты раздражающих ударов для перехода в тетанус, и Марей доказал, что разницы эти колеблются в довольно широких пределах (у людей): у одних одиночные сокращения, вызываемые отдельными индукционными ударами, совершаются сравнительно быстро, у других же медленно, и согласно с этим, для получения тетануса требуется применять на первых проворных М. большую частоту раздражений, нежели на вторых. Таким образом, индивидуальные разницы в степенях ловкости и проворства между людьми могут сильно зависеть от анатомо-физиологической организации их мышечной системы. Искусственными возбудителями М. могут служить свет и механические толчки, химические и термические раздражители. Раздражители последних двух родов вследствие длительности их действия могут давать только столбняки, тогда как механическими толчками можно вызывать как простые отдельные мышечные сокращения, так и тетанические, при действии целого ряда их (напр. колеблющимся молоточком тетаномотора Гейденгайна). Постоянный гальванический ток, пропускаемый через М., вызывает сокращение только в момент замыкания и размыкания тока, т. е. при появлении тока и исчезании его, или, другими словами, при внезапных колебаниях его силы, причем, при замыкании тока возбуждение исходит из отрицательного полюса, а при размыкании из положительного (Пфлюгер). Свет, прямо падающий на обнаженную М., при прерывистом мелькании его более 500 раз в секунду, вызывает также мышечное сокращение (д'Арсонваль). Искусственным раздражением М. человека электрическим током можно вызывать или простые мышечные сокращения, или столбняки; для первых прибегают к отдельным индукционным ударам, для вторых — к целому ряду их, быстро следующих друг за другом. Но при нормальных условиях М. возбуждаются нервными импульсами, приносимыми к ним по нервам из центральной нервной системы, и импульсы эти вызывают в них не простые одиночные сокращения, а тетанические. В пользу этого говорит 1) появление в сокращающихся М. при волевых или рефлекторных актах так наз. мышечного звука, тон которого соответствует около 19,5 колебаниям в секунду; в действительности же слышен гармонический звук основного тона, соответствующий 39 колебаниям в секунду; такой звук мыслим только при вибрациях мышечной массы, соответствующих отдельным простым мышечным сокращениям, слившимся в форму тетануса (Гельмгольтц); нет сомнения, что единичное мышечное сокращение не может вызывать вибрации М., а следовательно, и мышечного звука. Мышечный звук особенно удобно выслушивается при волевом сокращении жевательной М. (при полной тишине вокруг) или при сокращении двуглавой М. руки и выслушивании ее стетоскопом; 2) возникновение в М. ряда электрических колебаний (так назыв. отрицательных колебаний тока), из коих каждое отвечает простому единичному мышечному сокращению, приводящих в колебательное движение ртутный столбик в электрометре Липмана, а также и пластинку телефона, издающего при мышечном сокращении звук (конечно, в случае если эти аппараты были приведены в надлежащую связь с М.). Все нормальные мышечные сокращения являются поэт
| "БРОКГАУЗ И ЕФРОН" >> "М" >> "МЫ" |
Статья про "Мышцы" в словаре Брокгауза и Ефрона была прочитана 2938 раз
| Коптим скумбрию в коробке |
| Креветки с газировкой |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел