
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Корень, часть растений
Определение "Корень, часть растений" в словаре Брокгауза и Ефрона
Корень, часть растений (Radix). — Эта часть у большинства растений выражена весьма ясно и хорошо отличается от остальных, но немало и таких, которые или вовсе лишены К. или представляют переходы к стеблю и вообще обладают не типическими К. Не говоря уже о низших, слоевцовых растениях, как, напр. грибы и водоросли, мы находим и между цветковыми растениями такие, которые не представляют всех отличительных признаков типического К. В простейшем виде К. представляется у некоторых водорослей, состоящих из одной крупной клеточки, видимой простым глазом, напр. у каулерпы, у Botrydium и пр. Тут стеблеобразная часть клеточки выпускает вниз отростки, разветвляющиеся на множество тончайших мочек, как это бывает у первой из названных водорослей; или нижняя часть булавовидной клеточки, составляющей все растение, сама разветвляется наподобие корня. Но такие отростки служат только для прикрепления растения к почве и суть не что иное, как отпрыски единственной клеточки — это как бы подобие, а не настоящий К. Подобными же корнеобразными отростками, служащими для прикрепления к почве, снабжены крупные и даже гигантские морские водоросли. Мхи прикрепляются к почве многочисленными одноклеточными волосками, которые, вместе взятые, уподобляются пучковатым корням цветковых растений, но соответствуют скорее корневым волоскам высших растений (см. дальше), чем самим корням, — они по этому называются ризоидами. Настоящими типическими корнями снабжены собственно листостебельные растения. Уже в зародыше (см.) этих растений оконечность, противоположная его почечке, дающей начало стеблю, представляет собой К. в зачаточном состоянии. При прорастании она трогается раньше остальных частей зародыша и мало-помалу, вытягиваясь в сторону, противоположную направлению стебля, превращается в главный К. У многих растений, однако же, как у большинства однодольных, корешок зародыша рано замирает, а вместо него из основания стебля выступает несколько корешков, образующих вместе пучковатый К. Таким образом с самого начала все К. разделяются на 2 категории: главные и замещающие; многие, особенно деревянистые растения пускают, кроме того, из-под коры своих стволов или ветвей К. придаточные, появляющиеся в местах притока соков от повреждения или других причин. Так напр., К., выходящие из прутьев многих ив, тополей, смородин и пр., погруженных в воду, суть К. придаточные. Отличительные признаки типичного корня: он растет своей верхушкой, прикрытой особым чехликом (см. дальше); листьев никогда не приносит; он не имеет, за редкими исключениями (см. дальше), зеленого цвета, зависящего от хлорофилла; ветви его выступают из-под его коры — они эндогенные. Он может, однако же, пускать из себя стеблевые побеги, а потому и служит в некоторых случаях для размножения растения. Одни К. остаются на всю жизнь не ветвистыми, пуская только волоски или весьма тонкие веточки, такие К. называются стержневыми, напр. у моркови, у редьки, у свеклы; другие более или менее ветвятся, иногда до того, что главный ствол исчезает между ветвями, как, напр., у ели. По своей форме К. бывают конусообразные, репчатые, волокнистые и пр. По консистенции — мясистые, деревянистые и пр. Большинство растений пускает свои К. в землю, но у водяных плавучих они, очевидно, сами водяные, а у многих, селящихся на ветвях деревьев или вьющихся, как лианы (см.) вкруг стволов, имеются воздушные К. Таковы К. так называемых древожителей орхидных (см.) и ароидных (см.). Они отличаются от остальных тем, что заключают в себе хлорофилл. У некоторых растений К. представляются в виде коротких отростков, которыми они прицепляются к древесным стволам, к стенам, даже к гранитным скалам, как напр. у плющей. Точными опытами установлено, что главный К. направляется вниз по радиусу земли, но это вовсе не может служить общим правилом для корневых ветвей, которые нередко растут косо или горизонтально, у некоторых даже вверх. Несмотря на важность К., известно несколько растений, вовсе лишенных К. Таково, между прочим, орхидное Corallorhyza, y которого подземный, кораллообразно разветвленный стебель заменяет собой корень, выпуская только на оконечностях своих ветвей одноклеточные волоски.
А. Бекетов.
Анатомия корня. Анатомическое строение К, у многих растений существенно изменяется с возрастом. Различают первичное строение, свойственное всякому молодому К., и вторичное, свойственное более взрослым К. многих растений, но не всех. Первое особенно характерно для К., второе же весьма сходно с внутренним строением стебля. Сначала подлежит описанию первое, а потом второе. — Рассматривая поверхность молодого К., замечаем, что его верхушка (кончик) покрыта особым маленьким колпачком. Это — корневой чехлик. Обычно он бывает не больше нескольких мм и состоит из ткани более плотной, нежели прикрываемая им верхушка К. Отсюда понятна и роль его: он оберегает мягкий и нежный кончик К. от повреждений и поранений, в то время, как тот пробирается между угловатыми и острыми частицами почвы. У одних растений корневой чехлик остается навсегда, у других — сбрасывается в определенное время, когда становится далее бесполезным. На некотором расстоянии, как от верхушки, так и от основания К., поверхность последнего покрыта корневыми волосками. Волоски покрывают среднюю по возрасту часть К.: самая молодая часть еще без волосков, более старая (ближайшая к основанию) уже потеряла их. По мере того, как волоски физиологически изнашиваются и затем опадают, они заменяются новыми, молодыми, и такая смена идет по направлению от основания К. к его вершине или, как говорят, акропетально.
Фиг. 1. Ростки белой горчицы (Sinapis alba). А — с приставшими к корневым волоскам частицами почвы; В — после удаления последних через промывание ростка в воде.
Каждая клетка кожицы производит один волосок, так что волосок с произведшей его клеткой составляет одно целое — одну клетку. По форме — это длинные цилиндрические трубочки с закругленными концами, обычно достигающие длины в несколько мм. (0,1—8 мм.; фиг. 1 В) и тогда хорошо заметные простому глазу в виде нежных серебристых ниточек. Редко попадаются разветвленные волоски, еще реже многоклеточные. Правильную цилиндрическую форму волоски, впрочем, сохраняют только у К., развивающихся в воде или во влажном воздухе, тогда как у подземных В. (стало быть, у огромного большинства) они сильно изменяют свой облик, сообразно с теми мельчайшими полостями и ходами в почве, через которые им приходится пробираться; то они суживаются, то сильно расширяются. всячески изгибаются, плотно охватывают на пути мелкие частицы почвы, образуя при этом даже особые лопасти и тоненькие ворсинки, что дает им возможность еще более плотно примкнуть к этим частицам. Убедиться в существовании весьма энергичного сцепления между почвой и корневыми волосками можно посредством простого и наглядного опыта. Если молодые ростки, выращенные на песке [Мелкий песок часто употребляется как субстрат при искусственных культурах растений.], осторожно вытащим вместе с К. из почвы и затем осторожно отряхнем их, — тогда увидим, что песок легко опадет с молодой верхушки К. и с более старых его частей, т. е. как раз оттуда, где нет корневых волосков; там же, где они имеются, К. останется окутанным сплошным чехлом из песка. При дальнейших попытках отряхнуть песок вместе с песчинками отрывается уже много волосков. Понятно, что особенно хорошо такое срастание с частицами почвы видно при рассматривании волосков под микроскопом. Факт этот имеет большое физиологическое значение, о чем еще ниже будет упомянуто (физиология К.). Корневые волоски встречаются у огромного множества растений, тем не менее, не у всех. Так, не бывает их у подводных корней (Elodea, Lemna и др. водяные растения), у подземных корней многих хвойных и некоторых папоротников, также обычно у воздушных корней многих орхидных. Вообще присутствие или отсутствие их, большая или меньшая степень их развития определяются условиями жизни К., именно, главным образом, потребностью растения в воде и степенью влажности окружающей К. среды. Чем энергичнее растение испаряет воду, чем труднее, с другой стороны, достается она корням, тем более склонно оно к образованию обильных корневых волосков. Так, напр., у хвойных растений, испаряющих вообще мало воды, и корневые волоски развиты очень слабо или совершенно отсутствуют. У гиацинта или лилии волоски не образуются, когда К. растут в воде, и появляются в изобилии, когда корни растут во влажном воздухе. Отсюда видно, что развитием волосков можно до известной степени управлять по желанию. Интересно, что у некоторых растений (напр. у бука, дуба и др.) в обычных условиях корневые волоски не образуются, а их заменяют тоненькие гифы гриба, оплетающие молодой К. сплошным чехлом. Здесь гриб живет в симбиозе (см.) с К., составляя так наз. грибной К., микорицу, (Mycorhiza, см.). Поперечный разрез через молодой К. в области корневых волосков хорошо знакомит с внутренним устройством К. Прежде всего различаем: толстый периферический слой коры и тонкую центральную часть (центральный цилиндр Фан-Тигема), заключающую элементы сосудисто-волокнистого пучка (фиг. 2).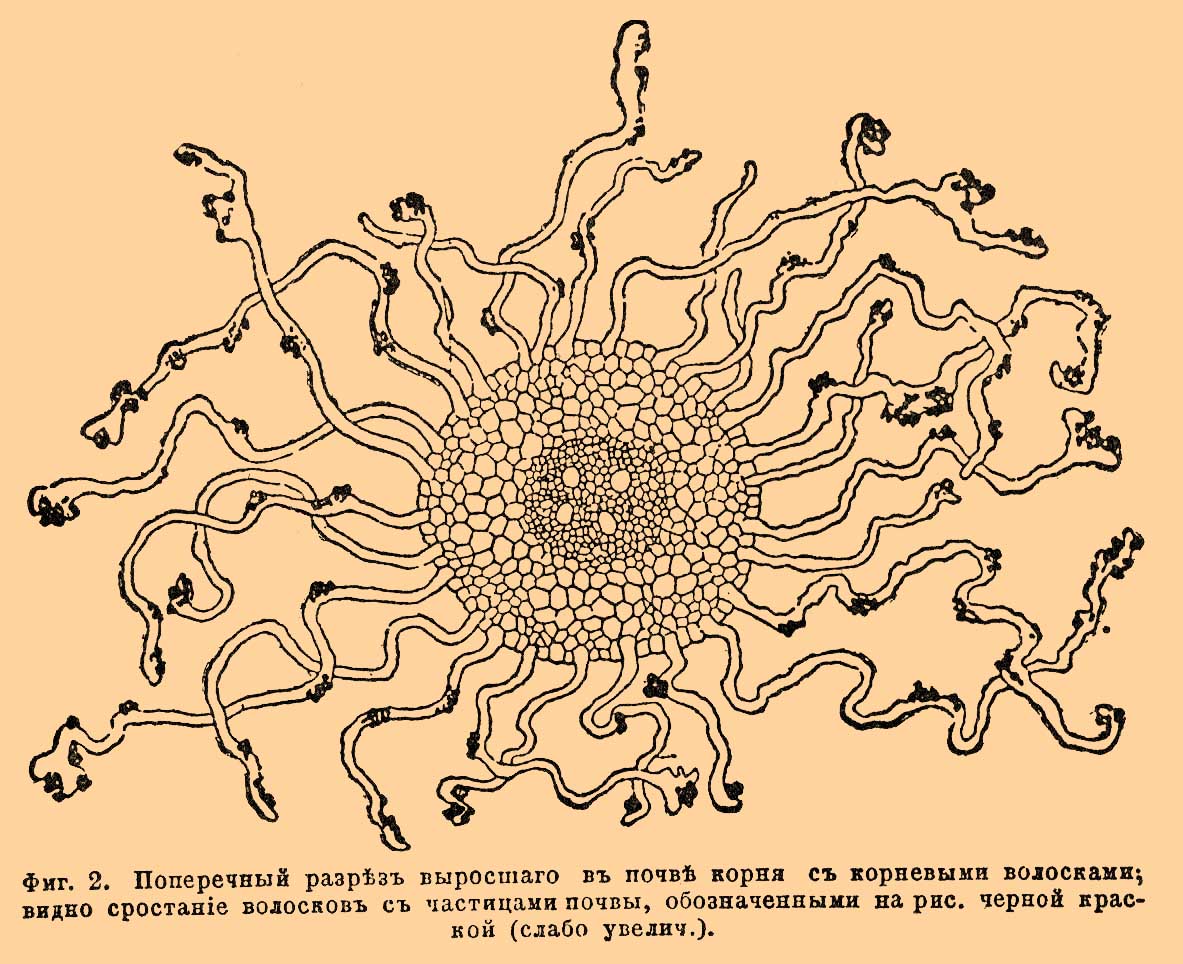
Фиг. 2. Поперечный разрез выросшего в почве корня с корневыми волосками; видно срастание волосков с частицами почвы, обозначенными на рисунке черной краской (слабо увелич.).
Кора состоит из тонкостенных паренхиматических клеток, располагающихся концентрическими слоями. Самый наружный ее слой составляет кожицу К. Клетки кожицы вытянуты в корневые волоски (фиг. 2) [В воздушных К. многих орхидных, некоторых Aroideae и др. растений кожица представляет особое любопытное уклонение в строении. Она не образует волосков, бывает не только однослойной, но и многослойной, и тогда превращается в особый довольно толстый покров К. (так наз. velamen radicis). Оболочки клеток такого покрова прозрачны или буроватого цвета, всегда кутинизированы и имеют, большей частью, спиральное или сетчатое утолщение. Сам покров является белым, губчатым или же буроватым, пергаментообразным. Нередко клетки покрова сообщаются отверстиями друг с другом и с окружающей средой. Внутри клеток нет живого содержимого и, смотря по степени влажности окружающей среды, они наполнены воздухом или водой. В первом случае покров приобретает серебристо-белый цвет, во втором через него просвечивает внутренняя зеленая (хлорофиллоносная) ткань. Покров способен жадно впитывать влагу и функционирует в качестве органа, снабжающего растение водой (особое приспособление к эпифитному образу жизни)]. Лежащий непосредственно под кожицей слой состоит из больших по величине многогранных клеток, вытянутых по радиусу более, нежели по окружности, и плотно сомкнутых своими боковыми стенками. По мере того как корневые волоски, а вместе с ними и клетки кожицы отмирают и опадают, оболочки этих клеток кутинизируются и весь слой приобретает свойства пробковой ткани, принимая на себя функцию наружного защитного слоя или экзодермы (внешняя эндодерма некоторых авторов). Таким же слоем и окутаны более старые части К., более не имеющие корневых волосков. Под экзодермой лежит более или менее толстый слой клеток, многогранной формы, плотно (без межклетных пространств) соединенных друг с другом, расположенных концентрическими слоями, но не образующих, однако, прямолинейных рядов по направлению радиуса. Размеры этих клеток возрастают от окружности к центру К., а развиваются слои в обратном направлении, т. е. центробежно. Это — наружная кора К. Вовнутрь от нее, до границы центрального цилиндра, идет внутренняя кора, слагающаяся из округлых или четырехугольных (на поперечном разрезе) клеток, правильно располагающихся концентрическими кольцами и в то же время радиальными рядами. В противоположность наружной коре, здесь клетки соединены между собой рыхло (образуются межклетные пространства, ср. фиг. 3 с); кроме того, их размеры уменьшаются постепенно снаружи внутрь, в том же направлении (центростремительно) идет и развитие слоев клеток.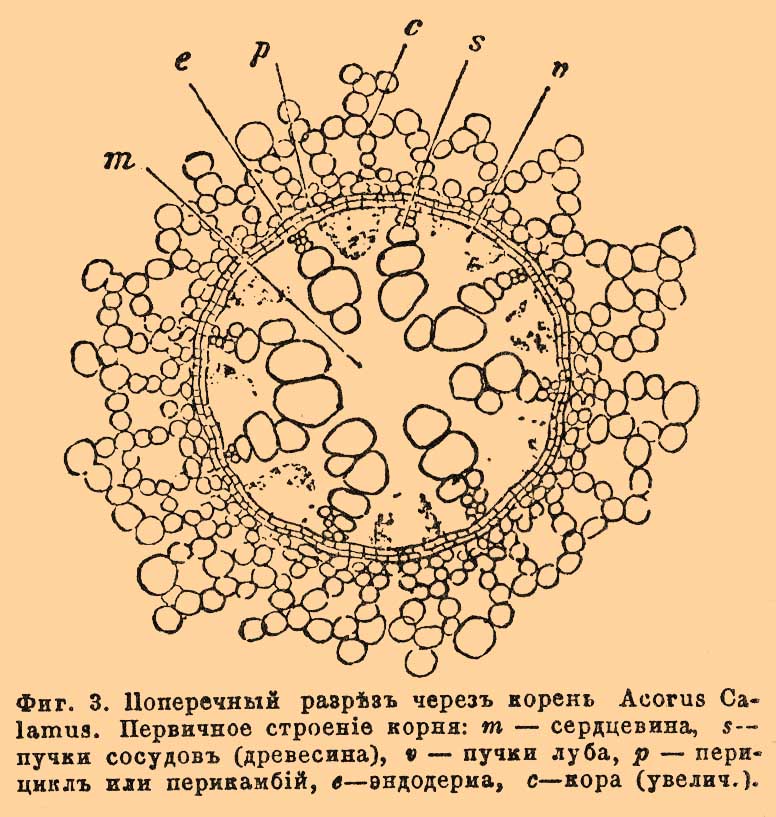
Фиг. 3. Поперечный разрез через корень Acorus Calamus. Первичное строение корня: m — сердцевина, s — пучки сосудов (древесина), v — пучки луба, p — перицикл или перикамбий, b — эндодерма, c — кора (увелич.).
Относительная толщина наружной и внутренней коры у различных растений весьма различна. Бывает (в очень тонких корнях ячменя [Hordeum], в корнях Lemna и др.), что наружная кора совершенно отсутствует, и наоборот — есть растения (Cycas, Epidendron и др.), у которых она достигает весьма значительной толщины, образуя собой почти всю кору К. У водяных и болотных растений внутренняя кора обычно очень толста. Межклетные пространства, находящиеся в наружной ее части, часто сильно увеличиваются и сливаются в широкие воздухоносные каналы, тянущиеся по всей длине К. Большие полости в этой части коры существуют также у злаков и у осок, но они иного происхождения. У воздушных и подводных К. наружная кора заключает много зерен хлорофилла. Самый внутренний и, вместе с тем, самый молодой слой коры слагается из клеток с кутинизированными оболочками и служит защитным влагалищем центрального цилиндра. Это внутренняя эндодерма или просто эндодерма (фиг. 3е, 4s, 5s); по своему строению ее клетки соответствуют вышеупомянутой экзодерме (подробнее в ст. Кора).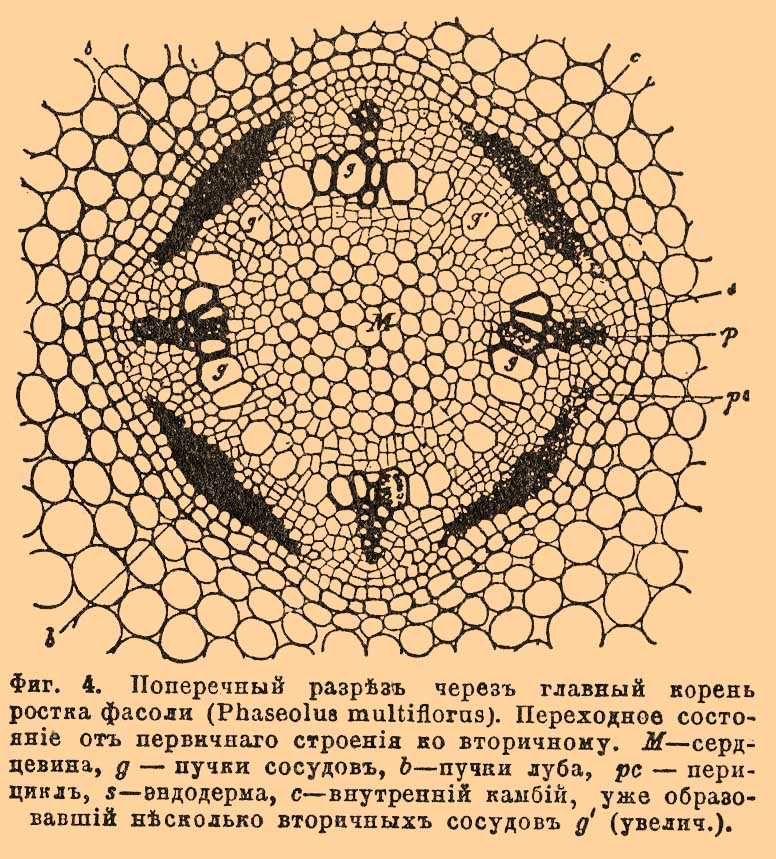
Фиг. 4. Поперечный разрез через главный корень ростка фасоли (Phaseolus multiflorus). Переходное состояние от первичного строения ко вторичному. M — сердцевина, g — пучки сосудов, b — пучки луба, pc — перицикл, s — эндодерма, c — внутренний камбий, уже образовавший несколько вторичных сосудов g' (увелич.).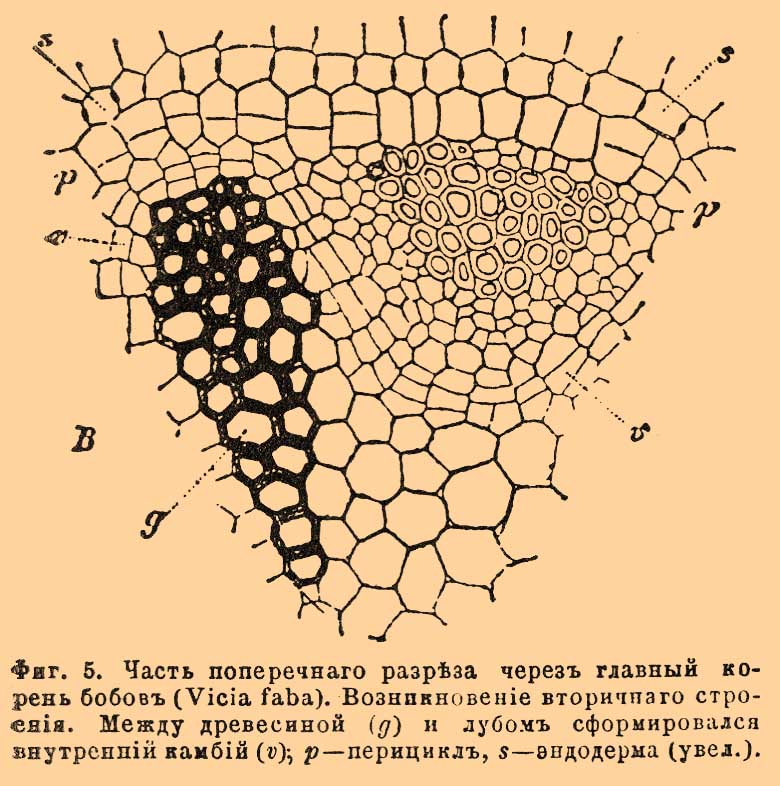
Фиг. 5. Часть поперечного разреза через главный корень бобов (Vicia faba). Возникновение вторичного строения. Между древесиной (g) и лубом сформировался внутренний камбий (v); p — перицикл, s — эндодерма (увел.).
Слой эндодермы в корнях образует хорошо заметную границу между областью коры и центральным цилиндром. Самый внутренний слой коры у сосудистых споровых растений является, кроме того, еще корнеродным слоем, т. е. из клеток его образуются боковые К. Центральный цилиндр начинается слоем нежных, тонкостенных, живых клеток, очень отличающихся от клеток эндодермы. Слой этот носит название перицикла или перикамбия (фиг. 3 p, 4pc); из его клеток возникают К. у цветковых растений. К клеткам перицикла, в двух или более, равно отстоящих друг от друга, точках прилегают радиально расположенные пучки сосудов (древесина), а в промежутках между ними залегают, также примыкая к перициклу, пучки лубяных элементов (фиг. 3 s, v; 4g, b) [Перицикл отсутствует у хвощей (Equisetum), пучки сосудов и луба примыкают у них прямо к удвоенной эндодерме. У многих злаков (рожь, овес, ячмень и др.), осоковых (Carex, Scirpus), ситовниковых (Juncus, Luzula) и у некоторых других семейств однодольных он прерывается против пучков сосудов, которые тогда упираются прямо в эндодерму. Перицикл, наоборот, прерывается против лубяных пучков, и тогда элементы луба подходят к эндодерме у Potamogeton, Najas и др. Попадается и многослойный перицикл; либо по всей окружности (Juglans, Pinus), либо против одних сосудистых пучков горох, бобы и др.) или, наоборот, — против одних лубяных (различные орхидные).]. Пучки древесины и луба, чередуясь, тянутся вдоль всего К. параллельно друг другу. Иногда древесинные пучки доходят до самого центра К. и здесь сталкиваются друг с другом. Таким образом, взаимное расположение древесины и луба в К. совершенно иное, чем в стебле: здесь нет таких сосудисто-волокнистых пучков, какие имеются там. Так как пучков сосудов и лубяных элементов бывает не менее двух каждого сорта и располагаются они симметрично, чередуясь, то и все строение К. является вполне симметричным по отношению к оси К. Де-Бари и другие ученые принимают весь центральный цилиндр за один сосудисто-волокнистый пучок, который и называют, по характерному расположению его составных частей, лучистым или радиальным. Пучки сосудов расширяются по направлению к центру К., — в этом направлении постепенно возрастает диаметр сосудов; а так как и само формирование сосудов происходит в центростремительном направлении, то, стало быть, самые узкие и самые периферические сосуды суть самые старые, а внутренние, наиболее широкие — самые молодые. Такой порядок образования сосудов весьма характерен для К. Узкие сосуды обычно бывают кольчатые или спиральные, тогда как широкие большей частью сетчатые, лестничные или точечные. Весь сосудистый пучок может низводиться у некоторых растений (водокрас [Hydrocharis], многие осоковые [Cyperaceae]) до 1 единственного узкого сосуда, — обычно же он содержит большее число сосудов, располагающихся в один или несколько радиальных рядов. У некоторых водяных растений (Alisma, Elodea, Vallisneria и др.) оболочки сосудов рано или поздно резорбируются, и на месте сосудов появляются воздухоносные полости и каналы. — В противоположность сосудистым пучкам, пучки луба шире к периферии, нежели к центру, к тому же они короче, т. е. не идут так далеко по направлению внутрь К. Они слагаются из ситовидных трубок, более широких во внутренней части пучка, более узких в наружной, и также образующихся центростремительно. Иногда пучок низводится до 1 ситовидной трубки — у Elodea, Potamogeton, 2—3 трубок — у пшеницы (Triticum), но обычно их в пучке гораздо больше. Число пучков сосудов и луба сильно варьирует, смотря по растениям и даже в одном и том же растении, смотря по величине К. и диаметру центрального тела. Оно не бывает менее 2 каждого сорта и может подыматься выше 100 (пальмы, Pandanus). Чаще всего бывает 2 — крестоцветные, маковые, гвоздичные, зонтичные, пасленовые, губоцветные, различные бобовые (Lupinus, Cytisus и др.), различные сложноцветные (Carduus, Cichorium), различные однодольные (напр., Allium), различные хвойные (напр., Cupressus): иногда 3 — Lathyrus, Vicia, Pisum и др.; часто 4 — просвирняковые, тыквенные и др. семейства; различные бобовые: Phaseolus и др., различные сложноцветные: Helianthus и др.; редко 5 — Faba, 6 — Quercus, 8 — Fagus. Впрочем, постоянство представляет только главный К., да и то не абсолютное, напр. у капуцина (Tropaeolum) бывает 2, бывает и 4 пучка, у хвойных (Pinus, Abies, Picea и др.) число варьирует от 3 до 14, смотря по видам и особям. У однодольных также нередко встречаются значительные колебания у различных особей одного и того же вида. — Первичное строение в корнях многих растений сохраняется на всю жизнь, подвергаясь с течением времени лишь весьма незначительным изменениям (склероз, кутинизация клеточных оболочек [Ткани молодого К. (имеющего первичное строение) образуются путем дифференцирования первичной меристемы верхушки К., меристема возникает, в свою очередь, через деление одной производящей (материнской) клетки или целой группы таких клеток (подробнее см. Ткани растительные).]). Так именно бывает у большинства сосудистых споровых растений, у большего числа однодольных и у некоторых двудольных (Ranunculus, Pinguicula, Trapa, Nympheaceae и др.). У остальных растений — у двудольных и у голосемянных — раньше или позже строение К. изменяется: К. приобретает новое строение — вторичное, — в противоположность первичному, очень сходное с анатомическим строением стебля. Обычно в К. образуется два деятельных слоя (2 камбия): внутренний и наружный. Первый возникает всегда в определенном месте. Сначала появляются камбиальные полоски, прилегающие к лубяным пучкам изнутри, а к сосудистом снаружи; потом они соединяются в непрерывный волнистый (на поперечном разрезе) слой. Получается камбиальное кольцо, функционирующее совершенно так же, как и в стебле: изнутри отлагаются древесинные, снаружи лубяные элементы, причем иногда образуются и сердцевинные лучи. Нередко можно также отличать и годичные слои, именно — вследствие различия между весенней и осенней древесиной. По своему строению корневая древесина сходна со стволовой, но рыхлее ее (так как состоит из более широких, но вместе с тем более тонкостенных элементов), а потому и не имеет ее технических достоинств. В противоположность внутреннему, наружный деятельный слой, так назыв. пробковый или перидермический камбий, может формироваться в самых разнообразных местах К., начиная от кожицы и кончая перициклом; чаще всего и у огромного большинства двудольных и голосемянных, он возникает именно из клеток перицикла. Снаружи этот слой порождает мертвую пробковую ткань, изнутри — живую феллодерму (см.). Все вместе (считая и сам камбий) зовется перидермой (см.). Когда перидермический камбий образуется из перицикла, то вся первичная кора скоро отмирает и сбрасывается в виде корки (см. Корка). Вследствие такой потери (остается один центральный цилиндр, покрытый тонким слоем пробки) К. сильно утончается, но потом он не только достигает прежней величины, но и утолщается постепенно, благодаря деятельности двух камбиев, особенно же внутреннего, вот почему у сосудистых споровых и у однодольных, у которых развивается один только наружный камбий, К. утолщается очень мало. В конце осени оба деятельные слоя перестают функционировать, остаются в покое зимой, а весной снова пробуждаются к деятельности. При всем сходстве К. со вторичным строением по внутреннему устройству со стеблем, не следует упускать из вида, что существует один особенно характерный для него признак, хорошо отличающий его анатомически от стебля — это положение первичной древесины в самом центре К.
Физиология К. Главных функций К. — 3: прикрепление растения к почве, поглощение воды и питательных веществ из почвы и препровождение их вверх в стебель. Положительный геотропизм главного К., вместе с явлениями циркумнутации, дают К. возможность проникнуть в почву и там укорениться. Энергия укоренения зависит от степени развития и расположения в почве корневой системы, от формы и глубины проникновения главного К., от числа рядов боковых К. и их расположения. Растение со стержневым К., как свекла или морковь, лучше укреплено в почве, при равенстве остальных условий, нежели растение с пучковатыми К., как пшеница или разные другие злаки; сосна с ее длинным, глубоко проникающим в землю К. лучше, нежели ель, К. которой не идут далеко вглубь. Укреплению растения в почве помогает еще способность К. укорачиваться в местах, уже переставших расти. С течением времени такое укорачивание может достигнуть 10% и даже 25% первоначальной длины К. В то время как молодая часть К. крепко сидит в почве, укорачивание старой части обусловливает все большее и большее втягивание в землю нижней части стебля, на которой затем появляются придаточные К., а это, в свою очередь, способствует еще большему укоренению растения. У некоторых пальм происходит, однако, явление противоположное. У пальм вообще главный К. замирает, а из основания ствола выступают толстые боковые К. Упираясь в почву, они выпирают ствол вверх, так, что его основание висит в воздухе, поднятое многочисленными К. Так бывает напр. у высокоствольной пальмы Iriartea exorrhiza и др. (см. Пальмы). Весьма интересно и важно, что К., укрепляющие растение в почве, в свою очередь, скрепляют саму почву. Этим пользуются для укрепления сыпучих песков. Чтобы остановить пагубное поступательное движение дюн, их засаживают растениями, способными жить на песках и быстро развивать обильно разветвленную корневую систему, таковы — Саrех arenaria, Elymus arenarius, Pinus maritima и некоторые другие [Относительно геотропизма, гелиотропизма, гидротропизма К. — см. эти сл. Воздушные К. многих растений из различных семейств (Orchideae, Aroideae, Bromeliaceae, Liliaceae и др.) обладают ясно выраженным отрицательным гелиотропизмом. То же встречается, но в гораздо слабейшей степени, и у некоторых подземных К., культивируемых в воде и подвергнутых одностороннему освещению (см. Гелиотропизм). Вообще же большинство подземных К. остается индифферентным к одностороннему действию света.]. Вторая функция К. состоит в поглощении из почвы воды и растворенных в ней веществ. Что растения всасывают воду К. — факт общеизвестный. Все знают, что после поливки почвы увядшие растения принимают свой прежний вид. При помощи довольно простых опытов (опыта Олерта), погружая К. в воду на различную глубину и, предохраняя оставшуюся на воздухе часть от засыхания слоем масла, наливаемого на воду, можно с несомненностью убедиться, что энергичное поглощение воды происходит почти исключительно в области корневых волосков. У растений, не образующих корневых волосков, дело не изменяется принципиально. И здесь всасывание происходит в области средней по возрасту, как раз соответствующей покрытой, притом, волосками у других растений. Зато в интенсивности процесса сказывается весьма существенная разница. Благодаря образованию длинных и тонких волосков, всасывающая поверхность К. чрезвычайно увеличивается — у гороха, напр., по исчислению Шварца, в 12 раз (см. любопытные указания в книге проф. Тимирязева), — а увеличение всасывающей поверхности для жизни растения имеет огромное значение. Из окружающего раствора К. не всасывает, наподобие губки или цедильной бумаги, без различия всякие вещества. Он берет то, что нужно для растения и оставляет ненужное или вредное [Впрочем, некоторые вещества, по-видимому, бесполезные, но зато и невредные, все-таки попадают в растение через К. Какие именно вещества всасываются К. из почвы, см. Минеральные вещества и Питание растений.]: он обладает особой избирательной способностью. Количество и качество всасываемого раствора находятся в зависимости от природы растения, кроме того, от его возраста и физиологического состояния. Вообще же: всосется корнем какое-либо вещество из почвенного раствора или нет — это зависит от способности вещества проникать через клеточную оболочку корневых волосков и постенный слой протоплазмы и от дальнейшей возможной судьбы вещества внутри растительных клеток [Подробнее см. в ст. Осмоз в растениях, Питание и Минеральные вещества растений.]. Старые, но поучительные опыты Соссюра хорошо иллюстрируют избирательную способность корня. Polygonum Persicaria, поглотив половину воды из данного ее К. раствора различных веществ, приняла растворенных в этой воде веществ не половину, а следующее число, в %: KaCl — 14,7, NH 4Cl — 12, Ca(NO3)2 — 4, Ca(C2H3O2)2 — 8, камеди — 9, сахара — 29, экстракта из удобренной почвы — 5. Стало быть, К. вбирали более воды, нежели растворенного в ней вещества и таким образом постепенно повышали концентрацию окружающего их раствора. Однако эти опыты Соссюра неприменимы прямо для суждения о поглощении К. веществ из почвы в обыкновенных нормальных условиях произрастания. Последующие исследования (а именно, опыты выращивания растений в искусственной культуре, в смеси питательных солей определенного состава) показали, что растворы Соссюра были чересчур концентрированными, тогда как наиболее пригодна гораздо слабейшая концентрация — именно 1—5:1000. Питательный раствор 1:1000 всасывается К. без изменения концентрации, а при еще меньшем содержании веществ в растворе поглощается уже больше растворенного вещества, чем воды, и раствор становится все менее и менее концентрированным. К. не только поглощает, но и выделяет из себя вещества, но явления последнего рода еще очень мало изучены. При помощи несложных опытов, однако, можно убедиться, что на поверхности К. находится кислая жидкость. Если, напр., К. развиваются во влажном воздухе, на поверхности синей лакмусовой бумажки, то, в местах прикосновения К., синий цвет лакмуса переходит в красный, даже каждый волосок оставляет после себя красный след. Другой опыт (Сакса) состоит в следующем. Мраморную полированную пластинку зарывают в горизонтальном положении в горшок с землей; в горшок сеют семена. Корни молодых растеньиц дорастают, наконец, до пластинки и затем ползут по ее поверхности. Если, спустя некоторое время, вырыть пластинку и затем внимательно рассматривать, то нетрудно заметить, что ее верхняя поверхность разъедена везде, где прикасались к ней К., представляя, таким образом, точный отпечаток К., со всеми его разветвлениями. Такое разъедание пластинки — результат действия на мрамор выделяемой К. кислоты. Не подлежит никакому сомнению, что К. выделяют углекислоту, ибо, как всякий живой орган, они дышат, т. е. поглощают кислород (O 2), и выделяют углекислоту (CO 2). В этом можно убедиться прямо опытами [Отношение объемов CO 2 и поглощаемого O 2 в равное время (CO2/O2) у К. всегда меньше 1, — напр. у К. Epilobium spicatum = 0,85. Согласно общему правилу (ср. Дыхание растений), более молодые части К. дышат интенсивнее, чем более старые.]. Выделяемая CO2 остается в газообразном виде, но чаще она растворяется в почвенной воде. Вода же, содержащая CO 2, растворяет углекислые соли кальция и магния; также и фосфаты растворимы в ней более, чем в чистой воде. Припомним теперь тот факт, что корневые волоски весьма плотно срастаются с частицами почвы. Такая близость весьма облегчает воздействие К. на нерастворимые в воде вещества. Благодаря выделению CO 2 и других кислот (разъедание мрамора), К. получает возможность питаться не только циркулирующим в почве раствором, но также утилизировать и твердые вещества почвы, переводя их предварительно в раствор. Огромное значение такого факта понятно само собой. — Третья главная функция К. состоит в препровождении поглощенных из почвы питательных веществ в стебель. Сильное осмотическое давление сока в клетках коры К. гонит поглощаемую корневыми волосками жидкость в сосуды древесины, сначала самого К., а потом и стебля. Нагнетательная способность К. носит специальное название корневой силы или корневого давления. Обусловливается ли она одним только осмотическим давлением сока в клетках паренхимы коры или же при этом имеют существенное значение и другие обстоятельства, вообще — причины корневой силы еще не вполне выяснены. Тем не менее, в существовании самой силы легко убедиться. Для опытов особенно пригодны растения с хорошо развитой корневой системой, из деревьев, например, береза или клен, далее виноградная лоза и многие травянистые растения — подсолнечник, георгина, крапива и др. Срежем у одного из таких растений стебель у самой земли; затем, обнажив К. на несколько сантиметров, укрепим на срезе, при помощи короткой соединительной трубки из каучука, длинную вертикальную стеклянную трубку. Выдавливаемый К. сок постепенно станет заполнять трубку, поднимаясь в ней все выше и выше, и, если почва достаточно влажная и теплая, поднятие будет продолжаться в течение нескольких дней (до 6—10 дней). При рассматривании среза К. в лупу, видно, что сок выступает из сосудов древесины. Если вместо простой трубки прикрепить к К. ртутный манометр, то можно определить силу давления, выталкивающего сок из К. ("давление К."; фиг. 6).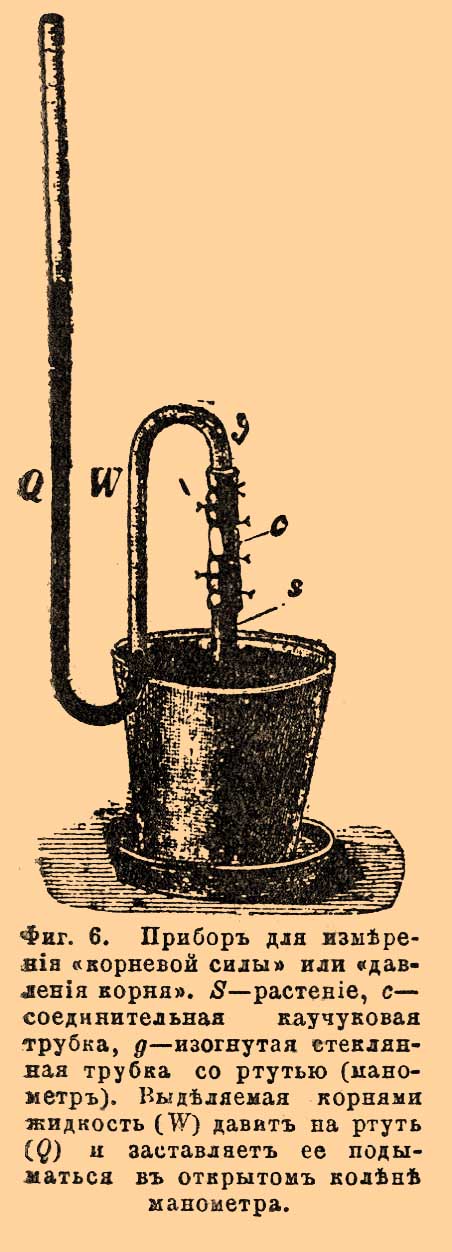
Фиг. 6. Прибор для измерения "корневой силы" или "давления корня". S — растение, c — соединительная каучуковая трубка, g — изогнутая стеклянная трубка со ртутью (манометр). Выделяемая корнями жидкость (W) давит на ртуть (Q) и заставляет ее подниматься в открытом колене манометра.
Выделяемый сок давит на ртуть и заставляет ее подниматься в открытом колене манометра. В опытах Гофмейстера давление сока уравновешивало колонну ртути у
| Phaseolus multiflorus | в 159 мм |
| Papaver somniferum | в 212 мм |
| Urtica urens | в 354 мм |
| Digitalis media | в 461 мм |
У виноградной лозы давление К. может превысить одну атмосферу. Так как вытекающий из разреза К. сок должен был ранее, при своем прохождении через всю длину К., преодолеть еще так назыв. внутреннее сопротивление К., то очевидно, что первоначальное осмотическое давление в клетках паренхимы коры еще значительнее. При помощи особых приспособлений (подробно см. Плач растений) можно также измерить и количество вытекающего сока. Если сопоставить объем вытекшего (за данное время) из К. сока с объемом самого К., то оказывается, что первый нередко в несколько раз больше второго. Вот данные опытов Гофмейстера:
| Продолжительность опыта | Название растения | Объем К. в мм 3 | Объем вытекшего сока, в мм 3 |
| 17—19 июня | Urtica urens | 1450 | 11260 |
| 4—18 мая | Urtica urens | 1350 | 3025 |
| 23—26 июля | Solanum nigrum | 1900 | 4275 |
| 26 июля—1 августа | Helianthus annuus | 3370 | 5830 |
Из таких сопоставлений и вышеизложенных опытов вполне выясняется роль К., как всасывающего и нагнетающего насоса. Выделяемый корнями многолетних растений сок неодинаков летом и весной. Летом он содержит главным образом минеральные вещества (минеральную пищу растения), и только следы органических: это почвенный раствор с очень незначительными примесями. Наоборот, весной сок содержит довольно много органических веществ, особенно сахара (весенняя пасока березы, клена и др.; см. Пасока). Выделение сока К. не происходит равномерно и всегда под одним и тем же давлением. В определенные часы дня количество выделяемого сока и корневая сила достигают максимальной величины, в другие часы опускаются до минимума. Изо дня в день можно наблюдать правильные колебания в высоте манометра, прикрепленного к К. Часы максимума и минимума у разных растений различны (подробн. см. Плач растений). — Когда растение испаряет больше воды, нежели К. успевает поглотить, то после удаления стебля К. не только не выделяет жидкости, но наоборот, еще сам способен ее поглощать. Действительно, если в жаркий летний день сделать вышеописанный опыт, то сок не только не будет вытекать из К. в трубку, но, даже если мы нальем в трубку воды, то она будет жадно поглощаться К. Лишь несколько часов спустя, к К. возвращается прежняя способность нагнетать сок в стебель. Интенсивное испарение воды листьями обусловливает разрежение воздуха в сосудах стебля и К., а это, в свою очередь, способствует поднятию сока из К. вверх в стебель и листья. Таким образом, сырой (восходящий) сок подвигается вверх благодаря толканию снизу (корневая сила) и тяге сверху (ср. Передвижение веществ в растении). Кроме 3 вышеописанных главных функций, К. несет еще некоторые второстепенные Так, он нередко служит складом для различных запасов питательных веществ (крахмала или сахара и т. п.), скопляющихся в клетках особой, сильно разросшейся паренхимы. Место образования такой паренхимы различно. Напр. в клубневидных К. орхидных это паренхима первичной коры, у Asphodelus — клетки сердцевины, у моркови — паренхима вторичного луба, а у репы или редьки — не одеревеневшая древесинная (древесная) паренхима, которая и составляет у них главную массу К. — Подробности и специальную литературу см. в следующих сочинениях: Бородин, "Курс анатомии растений" (СПб. 1888); Фаминцын, "Учебник физиологии растений" (1887); Van-Tieghem, "Trait é de botanique" (2 ed. 1 vol. 1891); Палладин, "Физиология растений" (2 изд. 1895); Тимирязева "Жизнь растения" (3 изд. 1895); Frank, "Lehrbuch der Botanik" (1 m. 1892); Strasburger, Schimper, Schenk und Noll, "Lehrbuch der Botanik" (1894).
Г. Надсон.
Корень (агроном. и лесов.). — Форма и большее или меньшее развитие К. обнаруживают существенное влияние на само развитие растений, выращивае
| "БРОКГАУЗ И ЕФРОН" >> "К" >> "КО" >> "КОР" >> "КОРЕ" |
Статья про "Корень, часть растений" в словаре Брокгауза и Ефрона была прочитана 4433 раз
| Коптим скумбрию в коробке |
| Салат тофу |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел