
БНБ
"БРОКГАУЗ И ЕФРОН" (121188)
- Photogallery
- Естественные науки - Математика - Технология
- Авиация и машиностроение
- Высокие технологии
- Вычислительная техника
- Нанотехнология
- Роботехника
- Энергетика
- Электроника
Грибы
Определение "Грибы" в словаре Брокгауза и Ефрона
Грибы (Fungi, Mycetes) — низшие растительные организмы, причисляемые к группе простейших споровых растений, к так называемым слоевцовым растениям. Все тело Г. слагается из очень тонких длинных нитей, называемых гифами [При нижеследующем изложении приняты во внимание только настоящие, гифенные грибы (Eumycetes auct., Hyphomycetes G.).]. Гифы могут быть одноклетными, без внутренних поперечных перегородок, или, что бывает гораздо чаще, внутри их замечаются поперечные перегородки, разделяющие гифу на отдельные клетки-членики. Часть тела, воспринимающая и перерабатывающая пищу, носит название грибницы, или мицелия. Размножение совершается помощью спор. Спора по своему назначению вполне соответствует семени высших растений, но сильно отличается от последнего по строению. Семя состоит из множества клеточек, даже заключает уже в себе зародыш, т. е. молодое недоразвитое растеньице, спора же почти всегда не что иное, как одна клеточка. Ближе всего к Г. по строению и циклу развития стоят некоторые водоросли. Но от них, равно как от всех других растений [О бактериях, водорослях без хлорофилла см. Водоросли и Бактерии. О бесхлорофилльных цветковых растениях сказано в ст. Сапрофиты и в других местах Словаря.], Г. существенно отличаются тем, что не содержат зеленого красящего вещества — хлорофилла. Отсутствие хлорофилла кладет отпечаток на весь склад жизни Г. Не имея возможности усваивать углерод из углекислоты, Г. добывают его из готовых уже органических соединений, следовательно, ведут сапрофитный, или чужеядный (паразитный), образ жизни. Распространенное в общежитии понятие о Г. весьма узко. Кроме съедобных Г. (см. отд. статью) и "поганок", наука относит к классу Г. еще множество и других организмов. Послание по большей части микроскопически малы, тем не менее значение их и для науки, и для практической жизни весьма велико. Об этом будет сказано ниже подробнее; кроме того см. Болезни растений, Головневые Г., Ржавчинные Г. и др. статьи. Переходим теперь к описанию отдельных частей и органов Г., их строению и развитию, к образу жизни Г., их значению в природе и для человека, их географическому распределению и к принципам их классификации.
ГРИБЫ I
Фиг. 1. Xylaria Hypoxylon. А — плодовые тела. В — поперечный разрез стромы с перитециями. С — перитеций с сумками (аскусами); а — сумка со спорами (300/1). D — образование конидий. Фиг. 2. Eurotium. А — мицелий (m) с конидиеносцем (с) и молодым перитецием (f) (490/1). В — отшнуровывание конидий. С — зачаток перитеция. D — молодой перитеций. Е — сумка со зрелыми спорами (600/1). Фиг. 3. Peziza calycina. A — в естественную величину. В и С — увеличено в 20 раз. Фиг. 4. А — нить капиллиция Bovista plumblea. В — базидии Geaster hygrometricus (390/1). Фиг. 5. Мицелий шампиньона с молодыми плодовыми телами. Фиг. 6. А — Гимениальный слой (200/1). В — базидии (500/1). Фиг. 7. Teichospora obducens. А — грибок в естественную величину (направо) и увеличенный в 20 раз (налево). В — сумки и парафиза (380/1). С — спермогоний. D — образование спермациа. Фиг. 8. Два вида хитридиевых грибков (а, а1), паразитирующие на водоросли. B — Chytridium Olla (400/1). Фиг. 9. Сапролегния с зооспорами (550/1). Фиг. 10. Сапролегния с половыми органами: антеридиями (а) и оогониями (А, В, С, D — оплодотворение. Е — ооспоры 550/1). Фиг. 11. Часть разреза через плодовое тело трюфеля: p — наружный покров, k — камеры с сумками (а), внутри сумок аскоспоры (200/1).
ГРИБЫ II
Объяснение к таблице II — см. в тексте.
Мицелий. В подходящих условиях споры Г. прорастают, образуя небольшую ростковую трубочку. На рис. 10 d и 5 е и f табл. "Грибы II" изображены различные стадии развития ростковой трубочки при прорастании спор обыкновенного строчка (Helvella esculenta). При достаточном питании ростковая трубочка вырастает у всех Г., за исключением Хитридиевых (Chytridieae, см.), где она остается рудиментарной, в более или менее длинную гифу. Гифы образуют обыкновенно много боковых ветвей, нарастающих и удлиняющихся, подобно главной гифе, своей верхушкой. На ветвях вырастают веточки и может получиться очень сложная разветвленная система гиф. Таким образом возникает и формируется мицелий. У целого ряда низших Г., у так называемых Phycomycetes (сюда принадлежат семейства Peronosporeae, Saprolegnieae, Mucorineae, Entomophthoreae и др.) мицелий остается, по крайней мере до стадии плодоношения, одноклетным. На рис. 1 табл. Г. II изображен такой мицелий, принадлежащий обыкновенной хлебной плесени Mucor Mucedo. У всех остальных Г. мицелий разгораживается на отдельные клеточки, становится септированным. Рис. 2 А табл. I представляет септированный мицелий плесневого грибка Eurotium repens. Мицелий служит для принятия и переработки (усвоения) пищи, во многих случаях также для хранения пищевых запасов, впоследствии утилизируемых организмом (напр., при образовании органов размножения). Мицелий может достигнуть значительного объема, являясь в виде ветвистых плотных шнурков, лент или плёнок, хорошо различаемых простым глазом. Таковы, напр., корневидные отроги, замечаемые у основания пенька (ножки) шампиньона и многих других шляпных Г., — это и есть остатки обильно разветвленного в почве мицелия (см. табл. I, рис. 5). Подобное мощное развитие мицелия особенно часто наблюдается у многих базидиальных Г. (Basidiomycetes). Обыкновенно, однако, мицелий весьма нежен, паутинообразен или хлопьевиден. Часто он совершенно не заметен простым глазом. Таков мицелий сапролегниевых, переноспоровых, головневых, ржавчинных и других Г. По образу жизни мицелий бывает сапрофитным и паразитным (чужеядным); сапрофитный — живет среди разлагающихся остатков животных или растений и питается отмершими органическими веществами; паразитный — гнездится среди живых тканей организмов и живет на их счет. Далее различают эндофитный чужеядный мицелий, живущий внутри питающего растения, и эпифитный, располагающийся по его поверхности. Весьма нередко только что указанные категории переносят и на целый организм Г. и говорят о паразитных, эндофитных Г. и т. д. У эндофитных Г. мицелий большею частью залегает в межклетных пространствах (интерцеллюлярно), но иногда он проникает и в самые клетки растения-кормильца (интрацеллюлярное распространение). Мицелии первого рода нередко дают с боку короткие отроги — сосала, проникающие внутрь соседних клеток. Отроги эти называются присосалами, или гаусториями (см.), и весьма типичны для некоторых родов, для Cystopus, напр., или для Peronospora. Обильно разветвленные гаустории Peronospora calotheca изображены на табл. II, рис. 9. У эпифитных мицелиев тоже образуются во многих случаях гаустории, внедряющиеся в клетки кожицы, но здесь они функционируют более в качестве присосок, удерживающих Г. на месте, чем в качестве органов принятия пищи (пример: виноградный Г. Erysiphe Oldium Tuckeri, его гаустории см. табл. Болезни растений).
Долговечность мицелиев весьма различна, от нескольких дней до многих лет. У очень многих Г. мицелий живет лишь определенный короткий срок, обыкновенно совпадающий с теплым временем года, а к осени отмирает; такой мицелий называют однолетним. Многолетние мицелии ежегодно приносят новые плоды. Однолетние, скоропреходящие мицелии типичны у головневых Г. (Ustilagineae), многолетние — у многих шляпных Г. и трутовиков (Hymenomycetes), также у некоторых паразитных ржавчинных Г. (Uredineae). Особый вид многолетнего мицелия представляет так называемый "покоящийся мицелий" (Dauermycel немецких ученых). Жизнь в нем как бы засыпает, питание и рост ослабляются и затихают. На стенах сырых погребов можно видеть темные мохнатые космы или покровы в виде плотного войлока — это и есть покоящийся мицелий. В прежнее время такие мицелиальные образования описывались под специальными родовыми названиями. Теперь же установлено несомненно, что это лишь стадии покоя других Г., по большей части гимениальных (Hymenomycetes). Так, напр., Rhizomorpha fragilis и Rh. subcorticalis входят в цикл развития обыкновенных опёнок. Весьма интересное изменение покоящегося мицелия представляют "склероции". Это более или менее округлые сферические или продолговатые тела в виде желваков или шишек, твердого, упругого сложения и темного цвета. Общеизвестные рожки спорыньи (Secale cornutum), попадающиеся на ржи, могут служить хорошим примером склероциев (см. фиг. 4 а, b, табл. "Болезни растений"). Склероции состоят из густо сплетенных и плотно скипевшихся друг с другом толстостенных гиф, образующих внутри склероция белую или светлоокрашенную сердцевину, а с периферии — темную кору (см. рис. 12, табл. II, а подробности строения в ст. Спорынья). Подобные склеротические образования входят как непременное звено в цикл развития самых различных Г., но для некоторых родов они особенно типичны. Среди дискомицетов — для рода Sclerotinia, среди пиреномицетов — для рода спорынья Claviceps, далее для многих гимениальных Г., напр., для обычного на навозе шляпного Г. Coprinus stercorarius. В тропических странах склероции достигают громадных размеров. Склероции, описывавшиеся прежде под названиями Pachyma и Mylitta и входящие, как оказалось впоследствии, в круг развития различных представителей родов Polyporus, Lentinus, Agaricus и др.,достигают величины кулака и больше, a Pachyma Cocos бывает часто величиной с голову. К этой же категории крупных склероциев принадлежит и склероций Polyporus tuberaster; он настолько тверд, что итальянцы прозвали его Pietra fungaja (грибной камень).
Спора. Споры суть отдельные клетки с протоплазматическим содержимым, клеточным ядром и оболочкой, которой нет лишь у зооспор. Как по строению, так и по величине они различны у разных Г. и даже нередко и у одного и того же Г. Все споры подводятся под несколько категорий. С морфолого-физиологической точки зрения различают подвижные споры (зооспоры, зоогонидии) и неподвижные. Зооспоры найдены сравнительно у немногих семейств Г. (Chytridieae, Ancylisteae, Saprolegnieae, Peronosporeae — все принадлежат к группе так назыв. Г.-водорослей [Phycomycetes], характеризующихся более простой, примитивной организацией). Это — голые клеточки (без оболочки), снабженные одной или двумя подвижными ресничками, при помощи которых они и движутся в воде, вращаясь или прыгая. В содержимом замечается, кроме клеточного ядра, пульсирующая вакуоль, а иногда еще и крупная капля масла (см. рис. двуреснитчатых зооспор на табл. I, 9 с и табл. II, 8 с). Неподвижные споры всегда обладают оболочкой. Оболочка может быть простой (одиночной) или двойной; в последнем случае различают внутреннюю оболочку, большею частью тонкую и бесцветную (эндоспорий), и наружную — более толстую, плотную, нередко цветную и украшенную рельефным узором (экзоспорий). Экзоспорий иногда утончен в тех пунктах, через которые впоследствии выйдут ростковые трубочки (Keimporen, поры прорастания). Однополостная (однокамерная) в начале клетка-спора у некоторых Г. разгораживается со временем перегородками на несколько отделений-камер. В этих многоклетных или многокамерных спорах каждая камера функционирует, как отдельная спора (см. табл. I, рис. 7 В).
По способу происхождения различают два сорта спор: экзогенные и эндогенные. Споры первого рода образуются поодиночке на концах гиф; обыкновенно их называют конидиями. Наиболее частый и распространенный способ образования конидий есть так назыв. отшнуровывание их, происходящее таким образом. На верхушке гифы (которая в таком случае называется конидиеносной, или "конидиеносцем", образуется вздутие, которое растет и превращается наконец в круглую или эллиптическую конидию; в то же время перешеек, соединяющий формирующуюся конидию с конидиеносной гифой, становится все уже и под конец совершенно прерывается. Образовавшаяся конидия или отваливается тотчас, или продолжает оставаться в соединении с произведшей ее гифой (см. рис. 2 В., табл. I). В последнем случае нередко под ней конидиеносная гифа образует новую, вторую конидию, под второй третью и т. д., таким путем получается целая "цепочка конидий", самая нижняя всегда самая молодая (рис. 7, табл. II). Устройство конидиеносцев и конидий будет рассмотрено при описании отдельных групп и семейств грибов. Нередко конидии образуются не непосредственно на концах конидиеносцев, а на верхушках особых выростков-веточек последних (стеригмы). Рис. 2 Ас. и В. табл. I представляет головчато-вздутую верхушку конидиеносца Eurotium, покрытую тесно стоящими кеглевидными стеригмами. Особую форму конидиеносцев, постоянную и типичную для некоторых групп грибов (Protobasidiomycetes, Basidiomycetes), представляет базидия. Стеригмы базидий различной длины и обыкновенно шиловидной формы. На верхушках их сидят конидии, называемые здесь базидиоспорами или просто спорами. Базидии могут быть цельными (одноклетными) или разделенными поперечными или продольными перегородками (о значении базидий для классификации Г. см. ниже; изображения базидий см. рис. 4 B и 6 B, табл. I). У большинства базидиальных грибов базидии тесно стоят друг около друга, образуя гимениальный слой, или гимений (Hymenium; рис. 6 А табл. I). Среди базидий в гимениальном слое часто рассеяны еще особые органы, с виду похожие на базидии, но всегда бесплодные (без стеригм и без спор, их называют парафизами и цистидами. Значение их еще неизвестно. Помимо базидий, и другого рода конидиеносцы могут собираться группами, образуя слои или пучки. Около таких групп может образоваться из гиф специальный покров и таким образом получится "конидиальный плод" (Conidienfrüchte). Конидиальные плоды, известные под именем "спермогониев" и "пикнид", составляют довольно типичную принадлежность некоторых групп грибов (Uredineae, Ascomycetes, см. рис. 1 D и 7 С и D, табл. I). Попав в подходящую питательную среду, конидии прорастают. Из конидий или вытягивается ростковая трубочка (подобно тому как из споры — молодая гифа), или из конидии выходят подвижные зоогонидии. Выхождение зоогонидий представлено на рис. 8 табл. II. Конидии могут размножаться еще почкованием. На одном или обоих концах конидии надуваются маленькие почки, постепенно достигающие величины самой конидии и в свою очередь дающие почки. Конидии обыкновенно остаются в соединении друг с другом, составляя цепочки различной длины. Подобный способ размножения конидий найден у многих грибов, он прекрасно наблюдается, напр., у головневых Г. (рис. 4, табл. II), но особенно типичен для дрожжевых грибков (Saccharomyces). По взгляду Брефельда, все тело дрожжевых Г. слагается из конидий; конидии обладают способностью беспредельно почковаться при благоприятных условиях, а гиф и настоящего мицелия не образуется никогда [Против такого взгляда имеется несколько веских данных и соображений, которые будут указаны в ст. Дрожжи.] (см. картину почкования дрожжей на рис. 6 табл. II).
Эндогенные споры образуются внутри клеток, чем и отличаются от экзогенных спор (конидий). Здесь опять две категории: либо спорообразование происходит внутри особых, специальных клеток, либо внутри обыкновенных вегетативных клеток мицелия. Эндогенные споры возникают или бесполо, или как результат полового акта. Клетки-вместилища, в которых развиваются (бесполо) споры, называются спорангиями. Строение и развитие их будет описано при обзоре отдельных групп Г. Здесь заметим только, что различают спорангии с неопределенным числом спор (собственно спорангии) и спорангии с ограниченным числом спор — обыкновенно 8, реже 2, 4, 16 и т. д., вообще кратным числа 2. Последние носят особое название аскусов, или сумок (Asci), и характеризуют собой очень большую группу Г., так и прозываемых сумчатыми Г., или аскомицетами (Ascomycetes). Сумки чаще всего булавовидной формы (рис. 1a, 2E, 7 В, табл. I), но бывают также овальные, эллиптические и др. (см. рис. 11, табл. I). Прежде полагали, что образование сумок со спорами является результатом полового процесса; однако этот взгляд мало-помалу оставляется [Подробнее см. Сумчатые Г.]. Сумки развиваются обыкновенно друг подле друга группами, образуя гимениальные слои и сумчатые "плоды" (перитеции, апотеции и др.). Несколько таких плодов представлено на табл. I (рис. 1c, 2F, 3 А- С; см. также Сумчатые Г., Пиреномицеты, Дискомицеты, Трюфель). Типичные спорангии с неопределенным числом спор попадаются у мукоров (Mucorineae). Содержимое крупного, видимого простым глазом шарообразного спорангия Mucor mucedo (рис. 1, табл. II — мицелий и спорангиеносная ветвь со спорангием; рис. 2 той же табл. — зрелый спорангий со спорами) распадается на множество отдельных эллиптических спор. Споры облечены оболочкой и высыпаются наружу после разрушения стенки спорангия. У других грибков, близко сродных мукорам, в спорангиях содержится меньшее количество спор, у Thamnidium Simplex (из той же группы Mucorineae) в спорангии всего одна спора. Такой спорангий, очевидно, морфологически весьма близок к конидии; в случаях же срастания оболочки единственной споры со стенкой спорангия (у некоторых Г.) спорангий становится совершенно эквивалентным конидии. Принимая во внимание еще и иные факты, приходят к выводу, что конидию нужно рассматривать как редуцированный спорангий. У Г., живущих в воде (Chytridieae, Saprolegnieae и др.), в спорангиях развиваются не неподвижные, покрытые оболочкой споры, а голые, снабженные ресничками зооспоры, по выхождении из спорангия активно уплывающие прочь. Спорангии с зооспорами называются зооспорангиями. Рис. 9, табл. I представляет два зооспорангия сапролегнии (водяная плесень): один зрелый, цельный, другой лопнувший, с вышедшими двуреснитчатыми зооспорами. Половой акт, ведущий к образованию спор, с полной достоверностью найден лишь у более просто организованных, низших Г. (Phycomycetes). Ближе всего он напоминает соответствующий процесс у водорослей: содержимое двух клеток, мужской и женской, сливается в одну массу, которая и становится спорой. Различают два типа оплодотворения и полового акта: один ведет к образованию ооспор, другой — зигоспор, или двоеспорий. При образовании ооспор внутри шаровидного женского органа (оогония), представляющего из себя одну клетку, из содержимого формируется одна (Peronosporeae) или несколько (Saprolegnieae) голых шаровидных масс (оосферы). Мужской оплодотворяющий орган (антеридий), имеющий вид боковой веточки, подходит к оогонию, и содержимое его через копуляционный канал вливается в оогоний, оплодотворяя оосферу или оосферы (стадии полового акта у сапролегнии изображены на рис. 10A-D табл. I). Оплодотворенные оосферы облекаются толстой оболочкой и превращаются в ооспоры (рис. 10Е, табл. I [такого рода половой акт носит название оогамии]. При образовании зигоспор две ветви мицелия, булавовидно вздутые, сталкиваются друг с другом; около места соприкосновения в той и другой ветви отделяется поперечной перегородкой по клеточке (рис. 3, табл. II, ветви — bb, клеточки — аа); перегородка между клеточками растворяется, и содержимое обеих клеточек сливается воедино, а потом окружается толстой крепкой оболочкой и таким путем превращается в зигоспору (двоеспорие, рис. 11, табл. II). Из сказанного и из рис. 3 видно, что разницы здесь между оплодотворяющей и оплодотворяемой клеточками нет никакой и нельзя сказать, какой элемент мужской и какой женский. Налицо тут лишь факт слияния; его принимают за простейшую форму полового акта и называют изогамией. Оплодотворение оосферы (яйца) подвижным сперматозоидом, столь распространенное среди водорослей и других растений, почти не встречается среди Г.; оно описано у одного только грибка Monoblepharis французским ученым Корню (Cornu). — Как упомянуто выше, эндогенные споры могут возникать и внутри обыкновенных вегетативных клеток мицелия. При этом содержимое клетки съеживается и покрывается новой, специальной, часто двойной оболочкой (головневые Г. и др.). В некоторых случаях содержимое собирается сначала к одному из концов клетки и здесь отгораживается поперечной перегородкой от остальной, опустевшей части клетки (Chlamydomucor). Возникшие таким путем споры носят название хламидоспор, немцы иногда называют их Gemmen — почками [К хламидоспорам близки так называемые оидии (oidien), считаемые некоторыми за конидиальную форму плодоношения. При образовании оидиев участок гифы сразу распадается на отдельные членики-споры.] .
Нередко один и тот же гриб размножается различными способами и приносит различные опоры. Так у сумчатых Г. рядом с типичными для них аскоспорами существуют еще конидии и хламидоспоры; у ржавчинных Г., кроме конидий, есть еще три сорта хламидоспор; у головневых Г. конидии и хламидоспоры, у гимениальных Г. — конидии (базидиоспоры), хламидоспоры и оидии, у пероноспоровых Г. — конидии и ооспоры и т. д. Различные формы плодоношения обыкновенно развиваются одна после другой, но бывают случаи, когда они появляются и одновременно, подчас даже на одном и том же участке мицелия. Это — случаи разноформенности или полиморфизма (плеоморфизма). О последовательности и правильности чередования отдельных форм размножения трудно сказать что-нибудь определенное. Бесполые поколения у низших Г. (Phycomycetes) могут сменить друг друга в неопределенном количестве прежде, чем наступит половое размножение, обыкновенно имеющее место в конце вегетативного периода. Равным образом аскоспоры и базидиоспоры часто появляются лишь под конец вегетации и завершают собой цикл развития индивида. Многие Г., с другой стороны, очень долго размножаются исключительно одними конидиями; есть немало и таких, у которых одна только эта форма размножения и известна, это так наз. Fungi imperfecti. Несомненно, что внешние условия влияют на появление того или другого способа размножения; но в чем это влияние — в большинстве случаев неизвестно. Во всяком случае у Г. не бывает правильного чередования поколений (Generationswechsel), всецело зависящего от внутренних причин, какое мы находим у высших споровых растений (мхи, папоротники). Чтобы составить себе верное и ясное представление о цикле развития какого-нибудь Г., особенное внимание нужно обращать на появление и смену различных форм размножения и плодоношения. Научные способы выращивания и воспитания Г. с целью уяснения их жизни и организации называются методами культуры грибов (см.). — В состав тела Г., подобно тому, как и у других организмов, входят минеральные (неорганические) и органические соединения. Из неорганических соединений первенствующее место занимает вода; некоторые сочные Г. содержат до 92,8% и более всего их веса в свежем состоянии (Agaricus melleus и др. шляпные Г.). Минеральные соединения, входящие в состав золы, не одинаковы у различных Г. и даже у одного и того же Г., в зависимости от почвы, среды. В золе каждого исследованного химически Г. были найдены: хлор, сера, фосфор, кремний, калий, натрий, кальций, магний и железо. Другие элементы попадаются в меньшем количестве и не всегда. Калий и фосфорная кислота явственно преобладают. Общее количество золы колеблется от долей процента до 15% всего сухого вещества. Из органических соединений первое место по количеству и важности занимают белковые вещества, углеводы и жиры (см. Грибы съедобные). Из углеводов у всех Г. найдена клетчатка (целлюлоза), в большинстве случаев в виде особой разности, так наз. грибной клетчатки, или фунгина, не дающего прямо (непосредственно) типичных реакций целлюлозы. Клетчатка является существеннейшею составною частью клеточных оболочек. Крахмала нигде не найдено; во многих случаях его замещает, по-видимому, гликоген. Сахары, слизи и др. углеводы встречаются также. Кроме названных органических соединений, попадаются еще: маннит, органические кислоты (особенно часто щавелевый кальций в виде мелких кристалликов на поверхности гиф), смолы, красящие вещества, ядовитые алкалоиды и много др.
Не обладая хлорофиллом, Г., как было упомянуто, не в состоянии питаться, подобно хлорофиллоносным растениям, одними только неорганическими соединениями. Питание (см. Питание растений) их в этом отношении сходно с питанием животных. Г., питающиеся разлагающимися (общее — мертвыми) веществами, называются гнилостными, или сапрофитами, живущие же за счет живых существ — чужеядными, или паразитами. И те, и другие играют значительную роль в природе. Сапрофиты — к ним принадлежит большинство Г. — возбуждают и содействуют (форсируют) процессам тления и разрушения мертвых растений и животных. Под влиянием жизнедеятельности этих Г. сложные органические соединения распадаются, сожигаются, окисляются. Продукты распада и окисления поступают в почву и воздух, а отсюда в том же или химически измененном виде воспринимаются хлорофиллоносными растениями, в которых снова перерабатываются в сложные соединения. Такова роль грибов в круговороте веществ в природе [Сами грибы, умирая и разрушаясь, удобряют почву для высших растений. См. также Симбиоз и Микориза.]. Г. и бактерии, стало быть, являются природными санитарами. Совсем иное нужно сказать о паразитных Г. Нападая на растения и на животных, не исключая и человека, они причиняют заболевания и повреждения. Грибные болезни животных и человека носят название микоз (mycosis) (см. Болезни растений, Растительные паразиты, Головневые, Ржавчинные Г., Парша, Молочница и др. спец. ст.). Заболевание может быть местным, локализированным, или общим, когда страдает весь организм. Г. болезни, оканчивающиеся смертью всего организма, не редкость. Болезни могут встречаться отдельными случаями — спорадически; но часто они принимают характер весьма губительных эпидемий (среди растений — головневые, ржавчинные Г., среди насекомых — энтомофторовые Г. и др.). Понятно, что такие паразиты сильно влияют на размножение видов (species), представители которых восприимчивы к заболеванию. Они являются в таких случаях не только тормозом, но до известной степени и регулятором развития вида. Самый механизм регуляции пока недостаточно ясен, но важность факта стоит вне всякого сомнения.
Человеку наиболее приносят пользу различные мясистые Г., идущие в пищу (см. Г. съедобные). В современной медицине употребление находит почти одна только спорынья (Secale cornutum — склероций грибка Claviceps purpurea) как средство, останавливающее кровотечения. В народной медицине на Западе до сих пор играют некоторую роль Auriculana Auricula ludae, Phallus impudicus, Elaphomyces cervinus и др.: другие виды (Lysurus, Torrubia) в большом ходу у китайцев. Гораздо важнее применение дрожжевых грибков (Saccharomyces cerevisiae и др. виды того же рода) в технике, именно, для приготовления алкоголя и различных спиртных напитков: вина, водки, пива, кумыса, кефира и др., также при хлебопечении (см. Дрожжи). В Японии для приготовления рисового вина (Sake) и некоторых кушаний с незапамятных времен применяется плесневый Г. Aspergillus Oryzae. В качестве полезных для человека Г. следует рассматривать некоторых паразитов, уничтожающих вредных насекомых, особенно вредных для лесов гусениц (Entomophthora, Empusa, Torrubia). Вред, приносимый грибами, может быть значительным. Некоторые Г., по виду схожие со съедобными, решительно ядовиты. Множество паразитных Г. нападают на разнообразнейшие возделываемые растения, причиняют им болезнь, иногда окончательную гибель, а хозяевам громадные убытки. Срубленные деревья (лес) и деревянные постройки подвергаются нападениям домового Г. (Merulius lacrimans), причиняющего "сухую гниль". Наконец, существуют и у человека болезни, несомненно вызываемые паразитными Г. (см. Растительные паразиты). О грибных болезнях шелковичного червя — см. Болезни шелковичного червя (т. IV).
Число известных видов грибов определяется проф. Саккардо (Saccardo, "Sylloge Fungorum etc.") в 32000. Если исключить виды, самостоятельность которых представляется весьма сомнительной, то в результате окажется, что число хорошо известных в настоящее время видов Г. никак не превышает 20000. Г. распространены по всей земле, повсюду, где возможна только растительность вообще. Теплый и влажный климат для них наиболее благоприятен, но они растут также и на далеком Севере, где найдено их около 1000 видов (Исландия 140, Шпицберген 70, северная Норвегия по ту сторону полярного круга 600). Наилучше известна грибная растительность умеренного пояса Европы и Северной Америки, который и представляется при современном состоянии сведений наиболее богатыми видами: для одной Германии приводится до 7000 в. Число тропических видов еще не может быть определено с точностью. Подробности географического распределения Г. мало известны, площадь обитания многих видов громадна, тем не менее допускают и для Г. существование географических областей, в общем совпадающих с таковыми же для цветоносных растений. Миграции из области в область тоже существуют. Относительно некоторых европейских Г., по крайней мере, несомненно доказано, что в Европу они попали лишь в самое последнее время (Puccinia Malvacearum, Plasmopara viticola). От Г., живших в прежние эпохи земли, осталось немного, что объясняется легкою разрушаемостью их тела. Следы их найдены в каменноугольном периоде; гораздо больше их известно из третичной эпохи (Hydnum antiquum Heer, Polyporites Bowmanni Lindl. и др.). Хорошо сохранившиеся мицелии, а иногда и зрелые грибки были находимы в янтаре. Руководящие принципы классификации Г. неодинаковы у различных ученых. Большинство думает, что водоросли, применяясь в течение многих поколений к сапрофитному или паразитному образу жизни, потеряли мало-помалу свой хлорофилл и превратились в Г. В пользу этого приводили как сходство строения, так особенно сходство форм полового акта [см . Водоросли.]. Сближая отдельные группы Г. с отдельными же группами водорослей (Saprolegnieae с Siphoneae, Mucorini с Conjugatae, Ascomycetes с Florideae и т. д.), склонялись к принятию полифилетического (т. е. от нескольких родоначальников) происхождения Г. В настоящее время ученые держатся преимущественно системы Брефельда (см. дальше); но и принципы классификации знаменитого Де-Бари не могут быть упущены из вида. Обращая главное внимание на способы размножения и особенно на половой процесс, он сопоставил отдельные группы Г. так:
I. Ряд сумчатых Г. (Phycomycetes)
1. Peronosporeae (рядом с ними Ancylisteae и Monoblepharis).
2. Saprolegnieae.
3. Mucorini, или Zygomycetes.
4. Entomophthoreae.
5. Ascomycetes.
6. Uredineae.
II. Группы, отклоняющиеся от ряда сумчатых Г. или с сомнительным положением в системе.
7. Chytridieae.
8. Protomyces и Ustilagineae.
9. Сомнительные Ascomycetes (Saccharomyces etc.).
10. Basidiomycetes.
Иную классификацию предложил О. Брефельд; основываясь на обширных и обстоятельных исследованиях, он пришел к выводу, что половой процесс встречается несомненно лишь у низших Г., так что форма полового размножения не может служить первенствующим классификационным признаком. Руководящие принципы у Брефельда: постепенное развитие мицелия, постепенное превращение спорангия сначала в несовершенную (Hemiasci), потом в типичную сумку (Ascomycetes), редуцирование спорангия до конидии, постепенная выработка из конидиеносца сначала несовершенной (Hemibasidii), потом совершенной, типичной базидии (Basidiomycetes) и наконец постепенное развитие и дифференцировка плодового тела (в отдельных группах) из простого однородного сплетения гиф. Из какой группы водорослей развились Г. — в точности неизвестно, принятие же происхождения (полифилетического) от нескольких групп по меньшей мере излишнее. Высшие Г., по Брефельду, ведут свое начало от мукоровых Г., Mucorini (Zygomycetes): сумчатые от спорангиеносных, базидиальные от конидиеносных. Промежуточными инстанциями являются Hemiasci и Hemibasidii. На прилагаемой таблице мы приводим основной план системы Брефельда (см. табл.).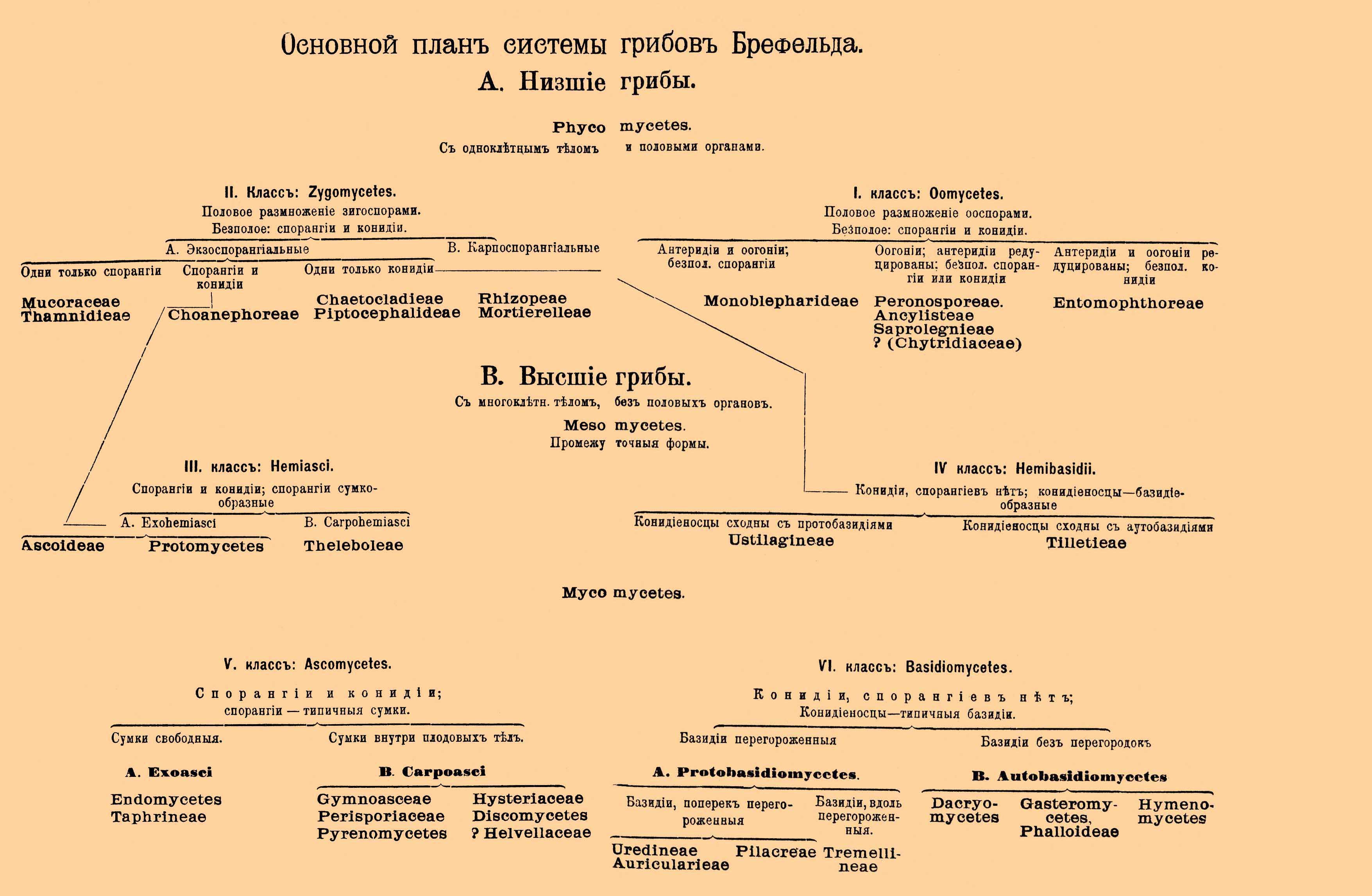
Существенно отличный от предыдущих взглядов на происхождение и систему Г. был развит проф. Гоби. Считать Г. редуцированными водорослями, по его мнению, нет никаких веских оснований. Г. произошли от амёбоидных организмов [См. ст. Миксомицеты, Монады.], стоящих на рубеже между растительным и животным царством. Амёбоидные организмы никогда не содержат хлорофилла и на вегетативной стадии (стадии роста и питания) представляются в виде голых комочков протоплазмы. Промежуточными формами между ними и настоящими грибами являются представители семейств Myxochytridieae [См. ст. Миксохитридиевые] и Reesieae. Ко взгляду профессора Гоби в последнее время присоединились и некоторые западноевропейские ученые (А. Фишер и др.). Окончательное установление естественного группирования Г. во всяком случае еще впереди.
Наука о грибах, микология, сформировалась в новейшее время. Первые научные исследования грибов относятся к началу прошлого 18-го столетия, но основателями новейшего направления следует считать Тюляна и Де-Бари, из коих первому обязана наука достановлением необыкновенно важного в общебиологическом отношении учения о полиморфизме, а второму — открытием половых органов у Г. Вся последующая деятельность микологов есть не что иное, как развитие того, что было открыто и установлено двумя названными великими ботаниками. Старое направление с преобладанием систематики и сухих описаний сменилось около сороковых годов нашего века новым, гораздо более плодотворным, выдвинувшим на первый план изучение организации и истории развития. Из новейших работ особого упоминания заслуживают обширные и важные исследования Брефельда. Литература микологии весьма богата; но здесь могут быть указаны лишь некоторые из наиболее важных сочинений, преимущественно общего содержания. L. В. Tulasne et A. C. Tulasne, "Fungi hypogaei" и "Selecta fungorum Carpologia" (1861-1865); A. De-Bary, "Vergleichende Morphologie und Biologie der Pilze, Mycetozoen und Bacterien" (Лпц. 1884); О. Brefeld, "Botanische Untersuchungen über Schimmelpilze" (1872, 1874, 1877); "Botanische Untersuchungen über Hefenpilze" (1880); "Untersuchungen aus dem Gesammtgebiete der Mykologie" (1887-91); L. Rabenhorst, "Kryptogamenflora von Deutschland, Oesterreich und der Schweiz" (2 изд.); J. Schröter "Pilze" (помещ. в A. Engler und K. Pranti "Die natürlichen Pflanzenfamilien", 1892), — W. Zopf, "Die Pilze" (1890); P. A. Saccardo, "Sylloge Fungorum omnium hucusque cognitorum" (I-Х, 1882-1892); Tavel, "Vergleichende Morphologie der Pilze" (1892). В этих сочинениях приводится остальная литература.
Г. Надсон.
| "БРОКГАУЗ И ЕФРОН" >> "Г" >> "ГР" >> "ГРИ" >> "ГРИБ" |
Статья про "Грибы" в словаре Брокгауза и Ефрона была прочитана 1883 раз
| Коптим скумбрию в коробке |
| Куриный суп |
TOP 15
- Волос
- Проно
- Степные животные
- Гимнастика
- Индийский океан
- Архитектура
- Сравнение, в литературе
- Манда
- Клитры
- Колесование
- Испарение
- Травоядные животные
- Оплодотворение у pacтений
- Вредные насекомые
- Электризация тел